Variability in heat tolerance among populations modifies the climate-driven periods of diurnal activity expected for ectotherm species. We illustrate this phenomenon for Iberian lizards in a paper we have just published in the Journal of Animal Ecology (blog post reproduced with permission by the Journal; see related blog).

Iberia is a wonderful natural laboratory, with a complex blend of flat/hilly, open/woody and coastal/continental terrain, swept by climatic gradients of temperature and moisture. In 2013, I launched a BES-supported project about the thermal ecology of Iberian lizards and managed to drive over much of the Iberian Peninsula in fairly little time. Not being a reptile specialist myself, I was confronted by the consistent observation that lizard populations occupied very different habitats across the known distribution of each of the ~ 25 known Iberian species belonging to the family Lacertidae.
For instance, the common wall lizard (Podarcis muralis) likes water, rocks and mountains, but you can find this pencil-long reptile at the top of a summit, along the slopes or riversides of shallow and deep ravines, on little stones barely surfacing above peatland grasslands, or among the bricks of buildings. These animals must experience different local climates conditional on where they live, and adapt their thermal physiology accordingly.
Having then started a postdoc in Miguel Araújo’s lab — a world-class site for global change ecology and ‘big’ biodiversity patterns — I reviewed a sizeable body of literature looking into large-scale gradients of thermal tolerance. Most of those papers had collated (mostly) one estimate of tolerance from each of tens to thousands of species, then mapped them against regional and global metrics of climate change through sophisticated mathematical frameworks. But these studies rarely accounted for population-level thermal tolerance.

At the time and in parallel with my project, the lab had just gathered a team of ecologists to do, to our knowledge, one of the largest one-off surveys of thermal tolerances in a group of closely related vertebrate ectotherms from a single biogeographic region in just two consecutive spring-summer seasons. This massive effort finally garnered upper and lower thermal limits (the highest and coldest temperatures an organism can withstand prior to death, also known as CTmax and CTmin, respectively) of 59 populations from 15 different species of lizards. We know that thermal traits are spatially and temporarily plastic — so much so for ectotherms (lizards in particular), whose body temperature relies heavily on air temperatures. So, we had the ideal data to assess to what extent such plasticity could shape our metrics of climate impact.
No sooner thought than done. The collation of the dataset involved driving over 25000 km for lizard sampling, allocating 200 hours to thermal assays, and 500 hours to lizard caring including terraria cleaning, and daily watering and life-prey feeding for each of 304 male lizards. We then statistically analysed the data in two steps with a focus on heat tolerance alone.
First, we found strong statistical support that single populations could not represent the heat tolerance of a full species, with (median) differences between populations of up to 3 °C. Such intraspecific variation nears the range of heat tolerance found across all of our study species! (median CTmax = 41.3 to 45.6 ºC).
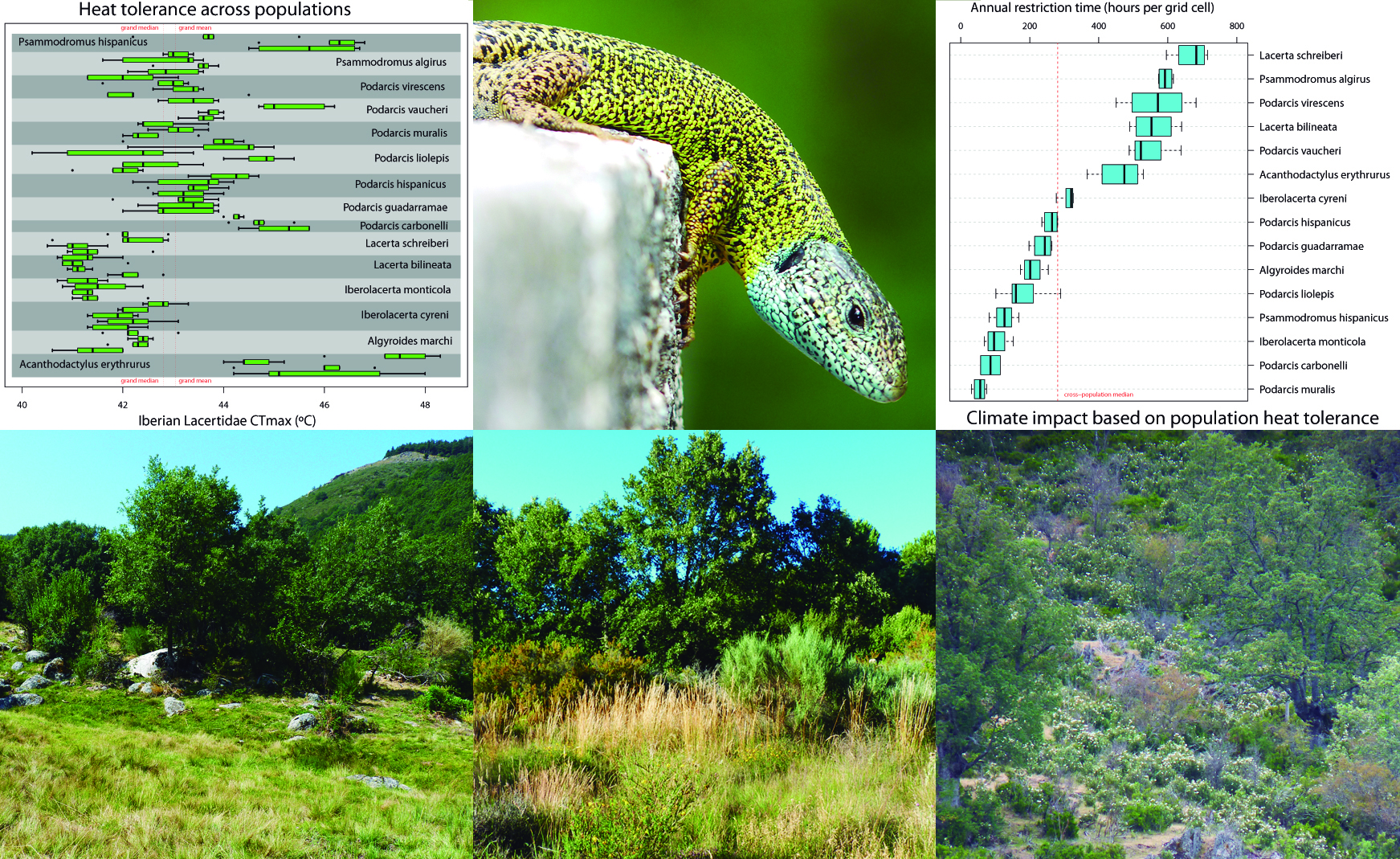
Second, we fed our population estimates of heat tolerance into a biophysical model encapsulating how an organism exchanges heat with the environment. We used this model to quantify the annual total of diurnal hours a species would be expected to be inactive (air temperature > CTmax) per unit of occupied area (25 km2). The rationale of our analyses was to mimic the widespread practice of choosing one population for a given species (see above), run our biophysical model to get an estimate of species-level restriction time, then repeat the same approach for each population sampled for that species.
We found that restriction times varied from 43 to 188 hours (median = 83 hours) depending on which population represented a species. For an average day with seven hours of sunlight, those discrepancies would imply intraspecific discrepancies in restriction time of 7 to 27 days (median = 11 days), and > 20 days for six of our study species. This is ecologically relevant because, during restriction times, lizards would be unable to feed or reproduce (unless accessing benign microhabitats not controlled for in our analysis). The take-home message pops up concisely in the title of our article: ‘Intraspecific variation in lizard heat tolerance alters estimates of climate impact’
In discussing our results, we were cautious about not falling into the straw-man fallacy that past macroecological studies using one thermal-trait estimate per species gets it wrong. Far from that. Previous research has revealed relevant patterns and posed critical hypotheses advancing the fields of macroecology and macrophysiology.
For example, many tropical species seem to live closer to their limits of heat tolerance and might be more sensitive to climate change than temperate species. Or heat tolerance appears to be less variable than cold tolerance, supporting the notion that the former might possess less scope to evolve in a warming scenario. Although we still lack a clear understanding of the evolutionary and adaptive drivers of such variation, our overarching argument is that controlling for intraspecific variation in thermal tolerance should improve our predictions of climate impacts on biodiversity.
Ecologists and physiologists have managed to gather information about the thermal ecology of just above 2000 species, but in most cases those species have been investigated at a single location each. Expanding taxonomic coverage is indeed a worthy effort to have a global picture of ecophysiological feedbacks under climate change – for planet Earth is teeming with at least several million species. Nonetheless, to address climate impacts in a comprehensively biological manner, future work should also endeavour to sample, map and model clines of thermal tolerance in the entire distribution of single species.
Salvador Herrando-PérezMore info:
Chown, SL, KJ Gaston. 2016. Macrophysiology – progress and prospects. Funct Ecol 30: 330-344
Herrando-Pérez, S, F Ferri‐Yáñez, C Monasterio, W Beukema, V Gomes, J Belliure, SL Chown, DR Vieites, MB Araújo. 2018. Intraspecific variation in lizard heat tolerance alters estimates of climate impact. J Anim Ecol doi:10.1111/1365-2656.12914
Valladares, F, et al. 2014. The effects of phenotypic plasticity and local adaptation on forecasts of species range shifts under climate change. Ecol Lett 17: 1351-1364





[…] the physiology of animals and plants and modify the magnitude of climate impacts (15) [see my blog here]. This phenomenon should be considered carefully in global ecology and to deal with global […]
LikeLike
[…] literature. Sometimes, I also write blogs to maximise the audience of my own publications (e.g., here and here), or to discuss a topic of general interest (e.g., numerical literacy). I have listed all […]
LikeLike