Climate change implies change in temperature and water, and both factors shape species’ tolerances to thermal stress. In our latest article, we show that lack of drinking water maximises differences in tolerance to high temperatures among populations of Iberian lizard species.

Climate change is a multidimensional phenomenon comprising temporal and spatial shifts in both temperature and precipitation (1). How we perceive climate change depends on whether we measure it as shift in (i) mean conditions (e.g., the mean air temperature or rainfall over a decade within a given territory), (ii) magnitude or frequency of extreme conditions (e.g., the frequency of floods or tornados or the number of days with temperatures or rainfall above or below a given threshold), or (iii) speed at which mean or extreme conditions change in space and/or time.
In aquatic ecosystems, climate change further alters water acidity, oxygen dissolution and melting of ice. However, many people, including some scientists, tend to equate climate change erroneously with increased mean temperatures. Psychologists have made the semantic point that the use of the expressions climate change and global warming as synonyms can give mixed messages to politicians, and society in general, about how serious and complex the climate emergency we are facing really is (2, 3) — see NASA’s simple-worded account on the subject here.
In our latest article (4), we reviewed the ecological literature to determine to what extent ecologists investigating the tolerance of terrestrial animals to high temperatures have looked at thermal effects over water effects. It turns out, they were five times more likely to examine temperature over water.
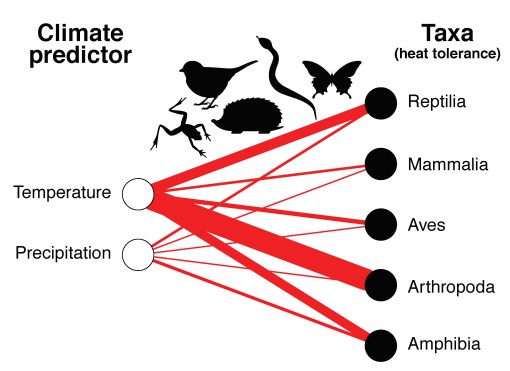
Frequency of correlations between climate (air temperature versus precipitation) and tolerance to high temperature of terrestrial fauna in 64 papers published in the ecological literature (thickest link = 36, thinnest link = 2) following a systematic literature review in Scopus (4).
This is counterintuitive. Just imagine you have been walking under the sun for several hours on one of those dog days of summer, and you are offered to choose between a sunshade or a bottle of water. I’d bet you’d choose the bottle of water.
The fact that body temperature and water balance are functionally integrated has been recently coined ‘thermo-hydroregulation’ (5). Essentially, if the environment in which an animal or human lives changes (e.g., increased temperatures, increased drought or both), they will have to deal with behavioural and physiological trade-offs in how hot (or cold) their bodies should get given available budgets of water.
This has been shown through experimentation using small lizards. For example, lizards prefer sun-exposed areas after storms, but shaded spots during dry periods (6), and the selection of hot, cold, wet or dry refuges is determined by environmental temperatures and water availability (7, 8). Consequently, thermal and water requirements jointly determine (micro)habitat selection and activity patterns, and therefore, biological responses to climate change (9) and extinction risk (10).
We have measured in prior studies that populations of Iberian lizards can differ by up to a median of to 3 ºC in the maximum temperatures they can tolerate across the distribution of individual species (11). Therefore, where exactly lizards of the same species occur in the Iberian Peninsula can tell a lot about how much heat they can take.
With that in mind, our study also examined if we could reproduce as much variation in thermal tolerance (3 ºC) by experimentally manipulating the availability of drinking water (4). We sampled two populations of each of two species of Iberian lizards: (i) the common wall lizard Podarcis muralis, a mesic species living in relatively high altitudes, and (ii) the spiny-footed lizard Acanthodactylus erythrurus, a xeric species widely distributed throughout Iberia.
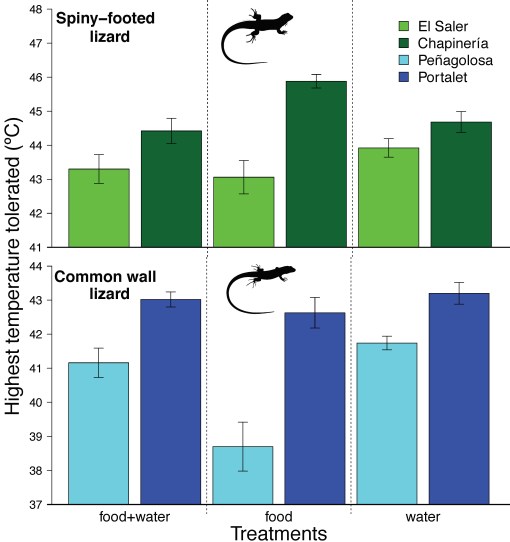
Maximum temperature tolerated by two populations* from two Iberian species of lacertid lizards. Species include the spiny-footed lizard Acanthodactylus erythrurus (green bars) and the common wall lizard Podarcis muralis (blue bars) in response to three experimental treatments (from left to right): provision of food and water, only food, and only water (4). Bars quantify mean CTmax ± standard error for 5 males/population/treatment. Lizard silhouettes by Ilian Velikov.
We found that water deprivation (combined with food supply) caused population differences in tolerance to high temperatures by 3 to 4 ºC; that is even larger population variation than measured in prior work (11). Those differences were two to three times greater if populations had to cope with food deprivation + water supply or with food + water supply.
This observation supports the prediction that, if faced with drought, and even if food is available, some populations will do far better than others in coping with extreme temperatures.
We were not surprised by our results, because we have often seen common wall lizards drinking from streams and puddles of water. In contrast, spiny-footed lizards might hardly see any water in many parts of their Iberian distribution (e.g., the sandy dunes of El Saler), but the fact that they drink water in experimental settings clearly indicates that exploiting such resources is part of their natural behaviour and imprinted in their evolutionary history.
Reptiles are sometimes associated with dry environments, but even species living in the driest regions of Earth will fancy a mouthful of water, hence the amusing observation that “… some desert lizards do drink when the opportunity arises from thundershowers” (12) — see video footage of reptiles avidly drinking here, here or here, or read here about the mesmerising strategy by which thorny devil lizards Moloch horridus can gather water from desert sands.

Iberian study sites (4). Spiny-footed lizards (left) were sampled at El Saler (Valencia) and Chapinería (Madrid) – top and bottom pics show a female and a juvenile with its red tail which honours the Spanish common name “colirroja”. Common wall lizards (right) were studied at Peñagolosa in my native province (Castellón) and Portalet with myself surrounded by the spectacular Pyrenees (Huesca) — both pics show males. All photos are mine.
This is all the more important for reptiles because we have known for decades that these animals obtain water through their diet (13, 14). However, dietary water is unlikely to replace the physiological role of drinking water (15, 16). Consequently, biological tactics for exploiting different sources of water must be major determinants of how species use their habitats and disperse through the landscape.
For instance, in Australia, the growth and body condition of the shingleback lizard Tiliqua rugosa can be best predicted if animals are modelled assuming that they have access to drinking water relative to dietary water and rainfall. In fact, the dry-continental margins of the distribution of this species are clearly correlated with water limitation (17). Across the board, ecologists must acknowledge that precipitation determines the magnitude of humidity and drinking water in an ecosystem, but those three factors along with dietary water can have different effects on the physiology of organisms (18, 19). To be biologically realistic, modelling patterns of species distributions and the processes that determine them should therefore discriminate the relative effects of different water resources.
Cold-blooded animals are currently facing two major threats. With global warming, they are gradually getting closer and closer to their upper limits of thermal tolerance (20), while their invertebrate prey are declining (21, 22). We contend however that the reduction in free-standing water brought about by increased aridification and drought (23, 24) can pose even a greater threat.
—
Acknowledgements: I shared most field and experimental work with Josabel Belliure while the project idea emerged from a brainstorming session with David Vieites. We are grateful to all co-authors for their contribution (Miguel Araújo, Wouter Beukema, Paco Ferri-Yáñez, John Terblanche and Thijs van den Burg), to Alicia Pérez Edo, Pedro Ballesteros, José Vicente Escobar and the University of Alcalá for logistic support, and to the British Ecological Society and the Spanish Ministry of Science and Innovation for funding. Permits for sampling and experiments granted by the following regional governments: Comunidad de Madrid (permit = 10/080485.9/16, Chapinería, Madrid), Gobierno de Aragón (INAGA/5000201/24/2016/3409, Portalet, Huesca), and Generalitat Valenciana (372/2015-VS(FAU15_038), El Saler, Valencia; and 2015/272-jvee, Peñagolosa, Castellón).
Overall, sampling lizards across Spain and Portugal has been the most enjoyable field-working experience of my entire career to date.
References
- Garcia, R.A. et al. (2014) Multiple dimensions of climate change and their implications for biodiversity. Science 344: 1247579
- Weber, E.U. (2010) What shapes perceptions of climate change? Wiley Interdisciplinary Reviews: Climate Change 1: 332-342
- Weber, E.U. (2016) What shapes perceptions of climate change? New research since 2010. Wiley Interdisciplinary Reviews: Climate Change 7: 125-134
- Herrando-Pérez, S. et al. (2020) Water deprivation drives intraspecific variability in lizard heat tolerance. Basic and Applied Ecology doi:10.1016/j.baae.2020.08.001
- Rozen-Rechels, D. et al. (2019) When water interacts with temperature: ecological and evolutionary implications of thermo-hydroregulation in terrestrial ectotherms. Ecology and Evolution 9: 10029-10043
- Ryan, M.J. et al. (2016) Too dry for lizards: short-term rainfall influence on lizard microhabitat use in an experimental rainfall manipulation within a piñon-juniper. Functional Ecology 30: 964-973
- Pintor, A.F.V. et al. (2016) Hydroregulation in a tropical dry-skinned ectotherm. Oecologia 182: 925-931
- Rozen-Rechels, D. et al. (2020) Water restriction induces behavioral fight but impairs thermoregulation in a dry-skinned ectotherm. Oikos 129: 572-584
- Pirtle, E.I. et al. (2019) Hydroregulation: a neglected behavioral response of lizards to climate change?, in Behavior of Lizards, V. Bels and other editors. (CRC Press, Taylor & Francis Group, Boca Raton, Florida, USA), pp. 343-374
- McCain, C.M. & Colwell, R.K. (2011) Assessing the threat to montane biodiversity from discordant shifts in temperature and precipitation in a changing climate. Ecology Letters 14: 1236-1245
- Herrando-Pérez, S. et al. (2019) Intraspecific variation in lizard heat tolerance alters estimates of climate impact. Journal of Animal Ecology 88: 247-257
- Munsey, L.D. (1972) Water loss in five species of lizards. Comparative Biochemistry and Physiology Part A: Physiology 43: 781-794
- Nagy, K.A. (1972) Water and electrolyte budgets of a free-living desert lizard, Sauromalus obesus. Journal of Comparative Physiology 79: 39-62
- Congdon, J.D. et al. (1979) Energetics, temperature and water relations in winter aggregated Sceloporus jarrovii (Sauria: Iguanidae). Ecology 60: 30-35
- Murphy, M.S. & Denardo, D.F. (2019) Rattlesnakes must drink: Meal consumption does not improve hydration state. Physiological and Biochemical Zoology 92: 381-385
- Wright, C.D. et al. (2013) Meal consumption is ineffective at maintaining or correcting water balance in a desert lizard, Heloderma suspectum. Journal of Experimental Biology 216: 1439-1447
- Kearney, M.R. et al. (2018) Field tests of a general ectotherm niche model show how water can limit lizard activity and distribution. Ecological Monographs 88: 672-693.
- Dupoué, A. et al. (2018) Reduction in baseline corticosterone secretion correlates with climate warming and drying across wild lizard populations. Journal of Animal Ecology 87: 1331-1341
- Lorenzon, P. et al. (1999) Effect of water constraint on growth rate, activity and body temperature of yearling common lizard (Lacerta vivipara). Oecologia 118: 423-430
- Sunday, J.M. et al. (2014) Thermal-safety margins and the necessity of thermoregulatory behavior across latitude and elevation. Proceedings of the National Academy of Sciences of the USA 111: 5610-5615
- Sánchez-Bayo, F. & Wyckhuys, K.A.G. (2019) Worldwide decline of the entomofauna: a review of its drivers. Biological Conservation 232: 8-27
- Lister, B.C. & Garcia, A. (2018) Climate-driven declines in arthropod abundance restructure a rainforest food web. Proceedings of the National Academy of Sciences of the USA 115: E10397-E10406
- Trenberth, K.E. et al. (2013) Global warming and changes in drought. Nature Climate Change 4: 17-22
- Park, C.-E. et al. (2018) Keeping global warming within 1.5 °C constrains emergence of aridification. Nature Climate Change 8: 70-74





[…] Basic and Applied Ecology1 — 10 years, 4 research papers2-5 [see related blog posts here, here, here and here], and 1 popular-science article6 after I joined the Department of Biogeography and Global […]
LikeLike