Just over two years ago I reported the ‘likely’ eradication of feral pigs from Australia’s third-largest (4,405 km2) island — Kangaroo Island. I indicated ‘likely’ because the program still required the proof-of-eradication phase to be completed before an official declaration could be made.
Yesterday I had the immense honour to take part in the official declaration ceremony held on Kangaroo Island near Karatta in the island’s southwest.
I can’t begin to emphasise enough how monumental this outcome is for biodiversity conservation, not just for South Australia, or even Australia, but globally.
This is without a doubt the largest island eradication of pigs anywhere in the world. Considering that most eradication (or even density-reduction) programs fail, the success is even more outstanding.
I am also taking some personal pride in the outcome because I had a small part to play. One dreary Sunday morning back in 2022, the head of PIRSA’s Invasive Species Unit Dr Brad Page rang me with an urgent request. “Corey”, he said without much preamble, “I need a model to tell me how much it would cost to get rid of pigs on Kangaroo Island, and I need it by this afternoon”.
Using animals as sport symbols reflects the integration of biodiversity into cultural identity and the transmission of collective values. This raises the possibility that the economic muscle of the sport industry could translate its symbolic capital into tangible commitments to biodiversity conservation.
Those who have had the privilege of travelling in remote areas might have come across an unexpected scene: a football pitch in the middle of the Amazon rain forest or on the slopes of the Andes, a basketball court on the side of a Buddhist temple, or an ice hockey rink on a snow-ploughed lake in remote northern Canada.
Sport is a global industry that generates identity, belonging, education, and shared emotions for both athletes and their avid spectators (1). Sporting affinities now rival the sense of nationhood once shared by citizens during warfare (2).
Now in our heavily monetised world, sport clubs rely on their fans through tickets and merchandising, and indirectly through television rights and advertising. In this both emotional and commercial relationship, expressions such as being true to the badge reflect the central role of corporate symbols in building bonds between a club and its supporters (3).
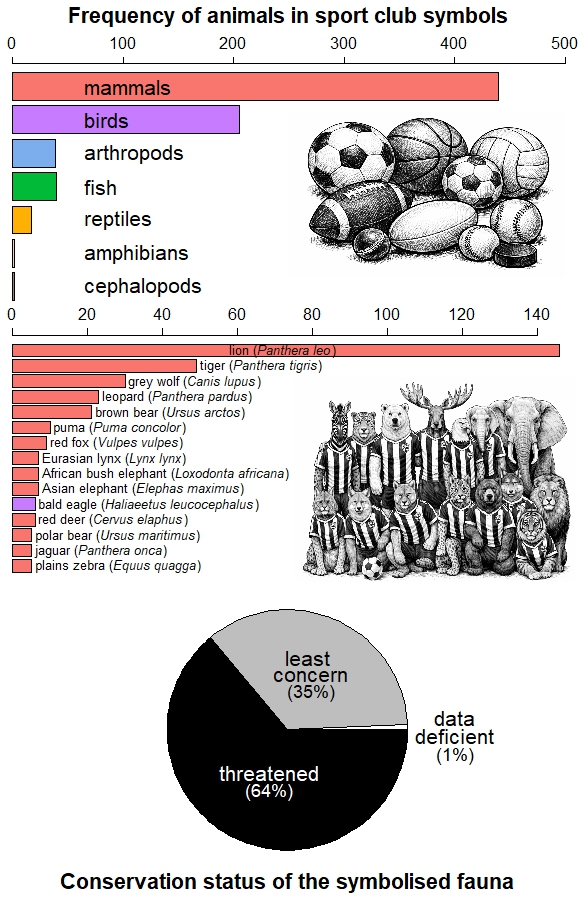
Sport club logos with animal iconography. Top row: examples of the grey wolf (Canis lupus) in Neftekhimik Nizhnekamsk (ice hockey, Russia), Warrington Wolves (rugby, England), Wolverhampton Wanderers (football, England), and Roma (football, Italy). Second row: bald eagle (Haliaeetus leucocephalus) in Essex Eagles (cricket, England), Adler Mannheim (handball, Germany), and Philadelphia Eagles (American football, USA). Third row: Free State Cheetahs (represented by the cheetah Acinonyx jubatus; rugby, South Africa), Toronto Blue Jays (blue jay Cyanocitta cristata; baseball, Canada), Memphis Grizzlies (grizzly bear Ursus arctos horribilis; basketball, USA) and Hisamitsu Springs (Japanese white-eye Zosterops japonicas; volleyball, Japan). Fourth row: six Spanish football clubs. Clubs featuring wolves and eagles are often associated with the symbolic qualities of these species (e.g., intelligence, prowess, fealty, bravery, strength). In football, animals reflect represent the history and heraldry of cities and regions, as seen in the crests of in Atlético de Madrid (brown bear Ursus arctos), AS Roma and Athletic Club (wolf), Cultural Deportiva Leonesa and Atlético Osasuna (lion Panthera leo), CD Castellón (raptor) and Levante UD (bat). Photos: Gary Kramer (wolf) and Andy Morffew (eagle).
In professional sport such as football, clubs increasingly function as brands (4) where even traditional logos are modified to enhance a team’s commercial value and strengthen audience loyalty (5). In this process, biodiversity becomes relevant because the iconography of many sport organisations incorporates representations of plants and animals.
Sport fauna
To quantify this phenomenon, Ugo Arbieu and collaborators analysed the presence of animals in club names, crests, and fan nicknames among 10 professional team sports across 50 countries (6). They found that 727 teams use 161 different animal species in their corporate imagery. Football and basketball lead in the number of species represented due to the large number of clubs worldwide, but American football, rugby and baseball display greater symbolic fauna diversity per club. Mammals and birds are the most common, particularly carnivores and raptors.
Animal symbols in club iconography (names, logos, fan nicknames) for the sports with the largest audiences (6): basketball, handball, baseball, cricket, football, American football, ice hockey and volleyball. The sample excludes 106 teams that use domesticated species as identity symbols, includes 163 men’s leagues and 67 women’s leagues, respectively, and the animal species depicted in the emblems of 48 teams could be identified. Horizontal bars above show the most represented animal groups (top panel), and the 15 species most frequently featured (middle panel). Bottom panel: percentage of symbols according to the IUCN’s conservation status of the species, where ‘threatened’ includes the categories Near Threatened, Endangered and Extinct. The trend indicates that sport clubs prefer to identify themselves with large mammal species that are threatened.
This pattern is not coincidental, for it reflects the historical bias of science and conservation towards large, charismatic vertebrates (7), but also the uneven availability of biological information and our social preferences for certain species (8). These preferences are even reflected in the animal emojis we share regularly on social media (9).
Arbieu’s study also revealed that clubs tend to favour images of threatened fauna (6), possibly due to their higher symbolic impact and media visibility (10). Moreover, although clubs in Europe and the Americas more often depict exotic animals, native species dominate in Africa, Asia, and Oceania (6). This suggests that the choice of an animal as an emblem is the product of not only aesthetic or symbolic criteria, but also of cultural roots and the historical relationship of societies with their local fauna.
Under the sea where there is little or no light, the foraging, communication, and orientation of whales and many other marine animals depend on sound. But increasing human activity has transformed the soundscape of seas and oceans. This change affects the behaviour of species and presents challenges in managing a problem of global scale.
Many like me feel uneasy when we hear a siren on the street. An ambulance, fire fighters, or the police can remind us of times when we or someone close to us suffered a heart attack, a fire, or a robbery. Animals can also associate sounds with risky situations they have experienced before, such as an attack from a predator, in their own lives or in the evolutionary history of their species.
For example, many types of whales are prey to killer whales (1) [watch predatory scenes here, here, here], and not only do they recognise the presence of their main predator by sound, but the vocalisations of some species have evolved to fall outside the killer whale’s hearing range (2). When faced with such a threatening sound, species must decide whether the risk of being hunted is great enough to justify interrupting essential activities such as feeding or mating (3). Interestingly, there are alarm signals that are so general in the animal kingdom, like a simple noise, that prey animals might react to them by spending time and energy to protect themselves, even when there is no real threat (4).
Tagging cetaceans off the Canary Islands to study their behaviour in relation to human and environmental disturbance. Above, 2 short-finned pilot whales (Globicephala macrorhynchus) in the southwest Tenerife, and below 2 Blainville’s beaked whales (Mesoplodon densirostris) in the Mar de las Calmas southwest of El Hierro. The back-mounted devices are DTAGs [read here, here and here]: they are attached by suction cups on top of which an encased electronic device records time series of environmental (depth, pressure, temperature, magnetic fields) and biological (e.g., swimming speed, heart rate, echolocation) variables. Watch videos of scientists deploying DTAGs on a range of cetacean species using a long stick here and here and drones here and here. Photos courtesy of O Marín Delgado (pilot whales) and C Yzoard (beaked whales); projects based at Universidad de La Laguna, Tenerife, Spain and led by N Aguilar de Soto [see stories here and here] (9, 26-29).
Naval sonar and killer whales
To examine this issue, Patrick Miller and his colleagues used underwater microphones to play recordings of killer whale sounds and ship sonar in the presence of 43 individuals from four cetacean species off the coasts of Norway and its Svalbard Archipelago (5): northern bottlenose whale (Hyperoodon ampullatus), humpback whale (Megaptera novaeangliae), long-finned pilot whale (Globicephala melas), and sperm whale (Physeter macrocephalus) [see press release for this research paper, listen to a podcast discussing findings]. During the experiment, each of the 43 individuals studied was fitted with a digital device attached to the skin using a suction cup. These devices recorded the animals’ movements and vocalisations. In total, the researchers collected 179 hours of baseline behaviour data in natural background noise, along with 7 hours of behavioural data in response to experimental playbacks of sonar [listen] and killer whale [listen] sounds.
Those of us living with cats share our homes with an ancestral predator, one adapted for hunting and the frequent, exclusive consumption of meat. These instincts become fully activated outside the domestic environment, where cats pose a global threat to wildlife.
Pets are family. We celebrate their arrival with the same joy as a grand homecoming, and their absence leaves a grief as deep as losing a loved one. In bonding with cats and dogs, we often attribute human abilities and emotions to them.
But beyond this affection, domestic animals still carry the instincts and genetic legacy of their wild ancestors(1, 2). My cats — Caruso, Muesli, and Plata — have been calm and loving, but they have always enjoyed a real hunt (3). When a moth comes in through a window, they seem possessed: their mouths chitter and make clicking sounds, they leap from one piece of furniture to another, and their heads snap sharply between the insect’s position and other points in the room, calculating the best spot from which to pounce on their prey. That is why when they become feral, cats and dogs integrate into food chains like any other species: they compete for ecosystem resources, hunt and are hunted, and hybridise and exchange diseases with other carnivores (4, 5).
Top: cat eating an Eurasian blue tit (Cyanistes caeruleus), a common visitor to home gardens in Nijmegen (Netherlands). Bottom, domestic cat after hunting an Eastern cottontail rabbit (Sylvilagus floridanus) in a residential neighbourhood of Stratford (Connecticut, USA). Photos courtesy of Jelger Herder (Nijmegen) and Scott Kruitbosch (Stratford). Scott is a photographer and conservationist. Near sunset on 30/09/2020, while intently observing local wildlife, he witnessed a neighbourhood cat sneak up from behind on a cottontail feeding in open grass and grab it. For years, Scott has had extremely negative interactions, both in person and online, with local residents over these issues. These exchanges have revealed that many people show little concern for wildlife or for the dangers their outdoor cats face, and believe that their cats would not, or could not, harm wildlife.
Domestic cats are highly skilled hunters, and their predatory interactions with a wide range of prey are widely documented in social media and documentaries. Some examples include cats catching: bats and birds on the wing, butterflies, chipmunks, dragons, fishes, grasshoppers, frogs, lizards, mice, owls, rabbits, seagulls, snakes, squirrels, and wallabies. See an award-winning photo depicting wildlife with fatal injuries caused by cats recorded in 2019 at a single animal hospital in the USA, and a video showing domestic cats mimicking bird calls and some cat owners explaining that their pets reject commercial cat food after experiencing the thrill of hunting real prey. The documentary Secret Life of Cats contextualises the ecological challenges posed by free-roaming cats.
Many animals avoid contact with people. In protected areas of the African savanna, mammals flee more intensely upon hearing human conversations than when they hear lions or sounds associated with hunting. This fear of humans affects how species use and move in their habitat.
Throughout our lives, we interact with hundreds of wildlife species without stopping to think about it. These interactions can be direct, such as encountering wild animals while hiking in the mountains or driving through rural areas — or more deliberate, as when we engage with wildlife for food, sport, or trade. As hunters, fishers, and collectors, we kill more than 15,000 species of vertebrates — one-third of known diversity — a range of prey 300 times greater than that of any other predator our size (1).
Now, let’s look at it from the other side. Anyone who has survived an attack or a fatal accident, they understand that the experience is remembered for a lifetime. Likewise, animals store information about threatening or harmful encounters with humans (2). For them, adjusting their behaviour in response to human presence has implications for their survival and reproduction (3, 4), which are passed down from generation to generation (5). This ability to adapt, for example, determines which individuals, populations and species coexist with us in urbanised environments (6).
Response to dangerous sounds
Liana Zanette and her team measured the flight responses of wild mammals in the Greater Kruger National Park (South Africa) when exposed to sounds that signal danger (7) [video-summary]. To do this, Zanette recorded videos of more than 4,000 visits to 21 waterholes by 18 mammal species. During each visit, a speaker attached to a tree randomly played one of five playback sounds: hunting dogs barking, gunshots, lion growls, human conversations in a calm tone and, as a control, the songs of harmless birds.
Deep-sea sharks include some of the longest-lived vertebrates known. The record holder is the Greenland shark, with a recently estimated maximum age of nearly 400 years. Their slow life cycle makes them vulnerable to fisheries.
In the Arctic, there are whales that have survived since the time of Napoleon’s Empire; in the Atlantic, there are molluscs that were contemporary with Christopher Columbus’ voyages; and in Antarctica, there are sponges born before the Holocene when humans were still an insignificant species of hunter-gatherers (see video on lifespan variation in wildlife).
Long-lived species grow slowly and reproduce at later ages (1, 2). As a result, these animals require a long time to form abundant populations and to recover from fishing-related mortality.
Among cartilaginous fish (chimaeras, rays, sharks, and skates), the risk of extinction due to overfishing is twice as high for deep-sea species compared to coastal species, because the former have longer and slower life cycles (3).
I used to think it was merely a post-COVID19 hiccough, but the extensive delays in receiving reviews for submitted manuscripts that I am seeing near constantly now are the symptoms of a much larger problem. That problem is, in a nutshell, how awfully journals are treating both authors and reviewers these days.
I regularly hear stories from editors handling my papers, as well as accounts from colleagues, about the ridiculous number of review requests they send with no response. It isn’t uncommon to hear that editors ask more than 50 people for a review (yes, you read that correctly), to no avail. Even when the submitting authors provide a list of potential reviewers, it doesn’t seem to help.
The ensuing delays in time to publication are really starting to hurt people, and the most common victims are early career researchers needing to build up their publication track records to secure grants and jobs. And the underhanded, dickhead tactic to reset the submission clock by calling a ‘major review’ a ‘rejection with opportunity to resubmit’ doesn’t fucking fool anyone. The ‘average time from submission to publication’ claimed by most journals is a boldface lie because of their surreptitious manipulation of handling statistics.
The most obese pachyderm in the room is, of course, the extortionary prices (and it is nothing short of extortion) charged for publishing in most academic journals these days. For example, I had to spend more than AU$17,000.00 to publish a single open-access paper in Nature Geoscience last year. That was just for one paper. Never again.
Anyone with even a vestigial understanding of economics feels utterly exploited when asked to review a paper for nothing. As far as I am aware, there isn’t a reputable journal out there that pays for peer reviews. As a whole, academics are up-to-fucking-here with this arrangement, so it should come as no surprise that editors are struggling to find reviewers.
In boreal forests, many hares adopt white winter coats before the snow arrives. In a snowless landscape, these white hares lack camouflage against predators. However, their early moult from brown into white fur can increase their survival and offers an advantage as the snow season becomes progressively shorter with climate change.
Throughout the year, we wear different clothing to protect ourselves from the cold or heat and for aesthetic reasons depending on the occasion. Likewise, many animals change the colour, thickness and structure of their fur and feathers in tune with the seasons.
Snowshoe hare (Lepus americanus) in a snowy (Kluane Lake/Yukon, Canada) and snowless habitat (Seely Lake/Montana, USA). This mammal moults its coat as colder temperatures, shorter days, and snowfall arrive. In the genetic populations of the temperate forests of the Rocky Mountains and the boreal forests spanning the North American continent, hares that moult from brown to white are abundant (20). However, in coastal areas, and in the third genetic population in the North Pacific, snowfall is brief and less intense, resulting in fewer white individuals. This is due to hybridisation with the black-tailed jackrabbit (Lepus californicus) over 3,000 years ago (17). The hare’s coat has an outer layer, where the longer fur gives each individual its colour, and an inner layer of short fur (19). In winter, the outer layer becomes thicker and denser, while the inner layer maintains a consistent thickness but increases in density. By biomass, the snowshoe hare is the primary herbivore in the North American boreal forest and distinguishes the trophic relationships between continents (21). In Europe, much of the boreal understory remains under snow, providing food for rodents with four-year abundance cycles controlled by small generalist predators (mustelids). In North America, the boreal understory grows above the snow and provides food for hares. In this region, snowshoe hare populations follow 10-year abundance cycles regulated by specialist predators (those that feed almost exclusively on hares), primarily the Canada lynx (Lynx canadensis) (6). Photos courtesy of Alice Kenney and Charles Krebs (Yukon) [see their ecological monitoring program here] and Marketa Zimova (Montana).
However, as the climate changes, springs arrive earlier, winters are delayed, and the frequency and intensity of precipitation have become highly variable. All of this makes it harder for species to adjust their wardrobe to temperature changes (1).
In this context, body colour is a critical factor for birds and mammals that undergo an annual moult (2). In 21 species from the cold latitudes of the Northern Hemisphere, some individuals are brown in summer, but turn white in winter, while others remain brown year round (3). This phenomenon includes weasels, rodents, ptarmigans, foxes, rabbits and hares.
Night is the peak activity period for many animal species. In the Western Andes of Ecuador, the Chocó golden scarab flies between forest patches during the night, but urban lighting interferes with their paths and jeopardises populations already struggling to persist in fragmented native forests.
Urban development has created a network of illuminated infrastructure that allows our society to function day and night without interruption. It is no surprise that with so much artificial light, we increasingly have to move farther away from towns and cities to see a sky full of stars.
Light pollution poses a challenge for nocturnal species that have adapted to living in the dimness of night (1, 2) — see documentaries about the impacts of artificial light on wildlife and insects, and a related scientific talk. This problem might be one of the causes of the global decline in insects (3, 4), in turn negatively affecting their role in maintaining agricultural systems through pest control, pollination, and soil quality (5). These concepts are featured by the documentaries The Insect Apocalypse and The Great Death of Insects.
Chocó golden scarab (Chrysina argenteola) walking on forest litter in La Maná (Cotopaxi, Ecuador). Growing to up to 4 cm in length, this species inhabits the tropical rainforest of the Chocó region in the Western Andes (10), where it is frequently attracted to artificial lights at night. The striking colour of this ‘jewel scarab’ is an optical illusion. The exoskeleton is covered with overlapping layers of chitin that polarise light and reflect hues of blue, gold, green, silver, or reddish tones, depending on the species (16). The metallic sheen appears to deter bird predation (17) and might serve as camouflage as well as aid in individual recognition (11). The eyes of insects are ‘compound’ — composed of 100s to 1000s of tubular eyelets (‘ommatidia’), each with its own cornea and lens (18), and all collectively contributing to insect vision. In nocturnal species like the golden scarab, the photoreceptor cells (at the base of each ommatidium) respond more slowly to light compared to diurnal species, allowing the former to collect more nocturnal light per unit of time before forming an image (19). However, just as staring at the sun blinds us, eyes adapted for night vision become overwhelmed by excessive artificial light, disrupting the behaviour of these species. Below the scarab image are two photographs contrasting the day and night landscapes of the same location in Pedro Vicente Maldonado (Pichincha, Ecuador) within the species’ distribution range. Photos courtesy of Martín Bustamante (animal) and Luis Camacho (city).
When flying, nocturnal insects orient their backs toward the sky, using the light of the moon and stars as a reference (6) (explained here and here). However, when they encounter artificial lights, they can no longer distinguish up from down, and so they can become disoriented, flying erratically, like a moth circling a streetlight.
It is estimated that a third of the insects attracted to artificial light die from collisions, burn injuries, exhaustion, and/or predation (7). In the tropics, finding countless dead insects at the base of urban lights is a common scene. Equally important is that artificial light also hinders migration, foraging, and the search for mates in many nocturnal species (1, 8, 9).
Nocturnal jewels
Camacho and collaborators evaluated the effect of artificial lighting at night on the Chocó golden scarab (Chrysina argenteola) (10). This species inhabits the tropical rainforests of the Western Andes from Ecuador to Colombia, and is a member of the group known as ‘jewel scarabs‘ due to their metallic body coloration (11). Because of its nocturnal habits and the larvae’s dependence on wood for food (12), the golden scarab has been increasingly affected by the loss of native forest in combination with light pollution from rural and urban expansion.
This is a fixed-term position for up to 3 years, and we are especially targeting Indigenous candidates.
The successful candidate will use existing code and develop new approaches to analyse complex data derived from lake, lagoon, river, and wetland cores measuring various aspects of dated vegetation composition, fire regime, and climate fluctuation. Additionally, the successful candidate will design simulation models to evaluate how different proxies behave under various environmental conditions, aiding in the interpretation of outputs from time-series models.
As a position under CIEHF, the position requires co-designing projects with Indigenous Partner Organisations, as well as extensive travel to the other Nodes within CIEHF to collaborate with palaeo-ecologists, climatologists, archaeologists, and other relevant specialists.
For more information and details on the application process, visit this link.
The internet has become an informational telescope to study what happens nearly everywhere the planet. Using internet observations, it has been recently documented that terrestrial hermit crabs use plastic waste as shelter along tropical coasts.
Before the internet irrupted, I was living in Spain and frequently travelled from my hometown to universities in Valencia and Barcelona to access scientific journals. Back then, these journals were only available in print or on compact discs. Today, I can do the same thing from home with an internet connection.
The emergence of public internet since the 1990s has globalised information and represents a data source for many areas of science (1, 2). When applied to nature, the term iEcology (internet Ecology) refers to the use of online documentation to study the natural history of plants and animals, their distributions, and the effects of humans on them (3). In fact, the internet highlights and promotes certain research topics. For example, bird species that are more frequently mentioned on social networks tend to be described taxonomically earlier, and are also the ones that interact most (positively or negatively) with human activity (4).
In search of the phenomenon
By exploring internet platforms Alamy, Flickr, Google, YouTube, and iNaturalist, Zuzanna Jagiello and her collaborators collected nearly 30 thousand photographs of hermit crabs to study the use of rubbish by these crustaceans (5). Hermit crabs are known for their peculiar habit of using empty snail shells to house their unprotected abdomens, carrying them around like someone travelling with their house on their back (6) — David Attenborough narrates here a funny swapping of shells among crabs of different size. The researchers aimed to assess the extent of the phenomenon of hermit crabs replacing natural shells with artificial materials as mobile homes (see video capturing the scene).
The Black Summer bushfires of 2019–2020 that razed more than half of the landscape on Kangaroo Island in South Australia left an indelible mark on the island’s unique native biodiversity, which is still struggling to recover.
Flinders Chase National Park on Kangaroo Island after the 2019-2020 Black Summer fires (credit: CJA Bradshaw)
However, one big bonus for the environment’s recovery is the likely eradication of feral pigs (Sus scrofa). Invasive feral pigs cause a wide range of environmental, economic and social damages. In Australia, feral pigs occupy about 40% of the mainland and offshore islands, with a total, yet highly uncertain, population size estimated in the millions.
Feral pigs are recognised as a key threatening process under the Environment Protection and Biodiversity Conservation Act 1999, with impacts on at least 148 nationally threatened species and eight threatened ecological communities. They are a declared invasive species and the subject to control programs in all Australian jurisdictions.
Motion sensing cameras deployed during the eradication program capture feral pigs using their snouts to search for soil-borne food. This behaviour, called rooting, creates large areas of disturbed soil, killing native vegetation and spreading invasive weeds and pathogens (credit: PIRSA).
Imagine growing up beside the eastern Mediterranean Sea 14,000 years ago. You’re an accomplished sailor of the small watercraft you and your fellow villagers make, and you live off both the sea and the land.
But times have been difficult — there just isn’t the same amount of game or fish around as when you were a child. Maybe it’s time to look elsewhere for food.
Now imagine going farther than ever before in your little boat, accompanied maybe by a few others, when suddenly you spot something on the horizon. Is that an island?
The western coast of Cyprus. CJA Bradshaw / Flinders University
When you beach your boat to have a look around, you can’t believe what you’re seeing — tiny boar-sized hippos and horse-sized elephants that look like babies to your eyes. There are so many of them, and you’re hungry after the long journey.
The diminutive beasts don’t seem to show any fear. You easily kill a few and preserve the meat as best you can for the long journey back.
When you get home, you are excited to let everyone in the village know what you’ve found. Soon enough, you organise a major expedition back to the island.
Of course, we’ll never know if this kind of scenario took place, but it’s a plausible story of how and when the first humans managed to get to Cyprus. It also illustrates how they might have quickly brought about the demise of the tiny hippopotamusPhanourios minor, as well as the dwarf elephantPalaeoloxodon cypriotes.
Non-native species introduced mainly via increasing trade of goods and services have huge economic, health, and environmental costs. These ‘biological invasions’ involve the intentional or unintentional transport and release of species beyond their native biogeographical ranges, facilitating their potential spread.
However, there is limited information available demonstrating whether a country’s capacity to manage its invasive species is effective at limiting future damage.
Our new study published in the journal Ecological Economics found that while more affluent countries with higher economic activity are vulnerable to more damage from invasive species, they also have the highest potential to limit damages incurred by investing more in management. Consequently, a nation’s economic capability partially determines the efficacy of investing in the control and prevention of invasive species.
In Australia, most fire occurs in the vast tropical savannas of the country’s north. In new research published in Nature Geoscience, we show Indigenous management of fire in these regions began at least 11,000 years ago – and possibly as long as 40,000 years ago.
But climate change and other effects of human activity are making wildfires more common and more severe in many regions, often with catastrophic results. In Australia, fires have caused major economic, environmental and personal losses, most recently in the south of the country.
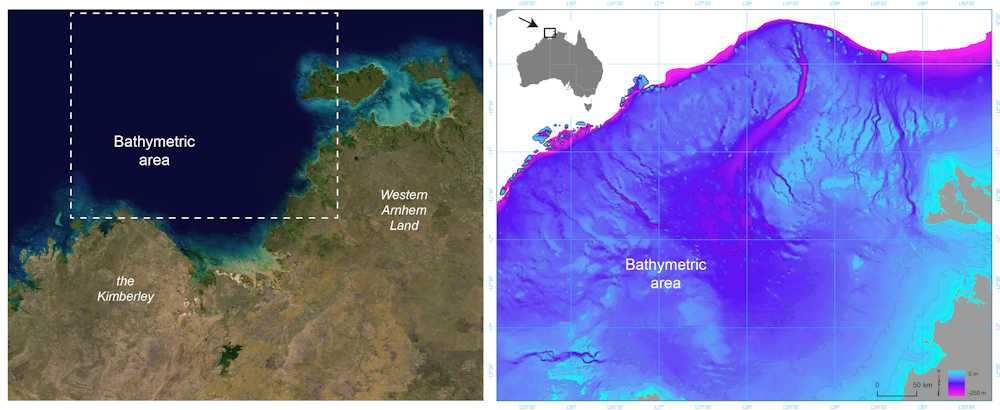
For much of the 65,000 years of Australia’s human history, the now-submerged northwest continental shelf connected the Kimberley and western Arnhem Land. This vast, habitable realm covered nearly 390,000 square kilometres, an area one-and-a-half times larger than New Zealand is today.
Left: Satellite image of the submerged northwest shelf region. Right: Drowned landscape map of the study area. US Geological Survey, Geoscience Australia
It was likely a single cultural zone, with similarities in ground stone-axe technology, styles of rock art, and languages found by archaeologists in the Kimberley and Arnhem Land.
There is plenty of archaeological evidence humans once lived on continental shelves – areas that are now submerged – all around the world. Such hard evidence has been retrieved from underwater sites in the North Sea, Baltic Sea and Mediterranean Sea, and along the coasts of North and South America, South Africa and Australia.
In a newly published study in Quaternary Science Reviews, we reveal details of the complex landscape that existed on the Northwest Shelf of Australia. It was unlike any landscape found on our continent today.
A continental split
Around 18,000 years ago, the last ice age ended. Subsequent warming caused sea levels to rise and drown huge areas of the world’s continents. This process split the supercontinent of Sahul into New Guinea and Australia, and cut Tasmania off from the mainland.
Unlike in the rest of the world, the now-drowned continental shelves of Australia were thought to be environmentally unproductive and little used by First Nations peoples.
But mounting archaeological evidence shows this assumption is incorrect. Many large islands off Australia’s coast – islands that once formed part of the continental shelves – show signs of occupation before sea levels rose.
Stone tools have also recently been found on the sea floor off the coast of the Pilbara region of Western Australia.
If several fossils of an extinct population or species are dated, we can estimate how long ago the extinction event took place. In our new paper, we describe CRIWM, a new method to estimate extinction time using times series of fossils whose ages have been measured by radiocarbon dating.And yes, there’s an R package — Rextinct — to go with that!
While the Earth seems to gather all the conditions for life to thrive, over 99.9% of all species that ever lived are extinct today. From a distance, pristine landscapes might look similar today and millennia ago: blue seas with rocky and sandy coasts and grasslands and mountain ranges watered by rivers and lakes and covered in grass, bush and trees.
But zooming in, the picture is quite different because species identities have never stopped changing — with ‘old’ species being slowly replaced by ‘new’ ones. Fortunately, much like the collection of books in the library summarises the history of literature, the fossilised remnants of extinct organisms represent an archive of the kinds of creatures that have ever lived. This fossil record can be used to determine when and why species disappear. In that context, measuring the age of fossils is a useful task for studying the history of biodiversity and its connections to the planet’s present.
In our new paper published in the journal Quaternary Geochronology (1), we describe CRIWM (calibration-resampled inverse-weighted McInerny), a statistical method to estimate extinction time using times series of fossils that have been dated using radiocarbon dating.
Why radiocarbon dating? Easy. It is the most accurate and precise chronometric method to date fossils younger than 50,000 to 55,000 years old (2, 3). This period covers the Holocene (last 11,700 years or so), and the last stretch of the late Pleistocene (~ 130,000 years ago to the Holocene), a crucial window of time witnessing the demise of Quaternary megafauna at a planetary scale (4) (see videos here, here and here), and the global spread of anatomically modern humans (us) ‘out of Africa’ (see here and here).
Why do we need a statistical method? Fossilisation (the process of body remains being preserved in the rock record) is rare and finding a fossil is so improbable that we need maths to control for the incompleteness of the fossil record and how this fossil record relates to the period of survival of an extinct species.
A brief introduction to radiocarbon dating
First, let’s revise the basics of radiocarbon dating (also explained here and here). This chronometric technique measures the age of carbon-rich organic materials — from shells and bones to the plant and animal components used to write an ancient Koran, make a wine vintage and paint La Mona Lisa and Neanderthal caves.
In light of new genetic research on the identity of ‘wild dogs’ and dingoes across Australia, the undersigned wish to express concern with current South Australia Government policy regarding the management and conservation of dingoes. Advanced DNA research on dingoes has demonstrated that dingo-dog hybridisation is much less common than thought, that most DNA tested dingoes had little domestic dog ancestry and that previous DNA testing incorrectly identified many dingoes as hybrids (Cairns et al. 2023). We have serious concerns about the threat current South Australian public policy poses to the survival of the ‘Big Desert’ dingo population found in Ngarkat Conservation Park and surrounding areas.
We urge the South Australian Government to:
Revoke the requirement that all landholders follow minimum baiting standards, including organic producers or those not experiencing stock predation. Specifically
Dingoes in Ngarkat Conservation park (Region 4) should not be destroyed or subjected to ground baiting and trapping every 3 months. The Ngarkat dingo population is a unique and isolated lineage of dingo that is threatened by inbreeding and low genetic diversity. Dingoes are a native species and all native species should be protected inside national parks and conservation areas.
Landholders should not be required to carry out ground baiting on land if there is no livestock predation occurring. Furthermore, landholders should be supported to adopt non-lethal tools and strategies to mitigate the risk of livestock predation including the use of livestock guardian animals, which are generally incompatible with ground and aerial 1080 baiting.
Revoke permission for aerial baiting of dingoes (incorrectly called “wild dogs”) in all Natural Resource Management regions – including within national parks. Native animals should be protected in national parks and conservation areas.
Cease the use of inappropriate and misleading language to label dingoes as “wild dogs”. Continued use of the term “wild dogs” is not culturally respectful to First Nations peoples and is not evidence-based.
Proactively engage with First Nations peoples regarding the management of culturally significant species like dingoes. For example, the Wotjobaluk nation should be included in consultation regarding the management of dingoes in Ngarkat Conservation Park.
Changes in South Australia public policy are justified based on genetic research by Cairns et al. (2023) that overturns previous misconceptions about the genetic status of dingoes. It demonstrates:
Most “wild dogs” DNA tested in arid and remote parts of Australia were dingoes with no evidence of dog ancestry. There is strong evidence that dingo-dog hybridisation is uncommon, with firstcross dingo-dog hybrids and feral dogs rarely being observed in the wild. In Ngarkat Conservation park none of DNA tested animals had evidence of domestic dog ancestry, all were ‘pure’ dingoes.
Previous DNA testing methods misidentified pure dingoes as being mixed. All previous genetic surveys of wild dingo populations used a limited 23-marker DNA test. This is the method currently used by NSW Department of Primary Industries, which DNA tests samples from NSW Local Land Services, National Parks and Wildlife Service, and other state government agencies. Comparisons of DNA testing methods find that the 23-marker DNA test frequently misidentified animals as dingo-dog hybrids. Existing knowledge of dingo ancestry across South Australia, particularly from Ngarkat Conservation park is incorrect; policy needs to be based on updated genetic surveys.
There are multiple dingo populations in Australia. High-density genomic data identified more than four wild dingo populations in Australia. In South Australia there are at least two dingo populations present: West and Big Desert. The West dingo population was observed in northern South Australia, but also extends south of the dingo fence. The Big Desert population extends from Ngarkat Conservation park in South Australia into the Big Desert and Wyperfield region of Victoria.
The Ngarkat Dingo population is threatened by low genetic variability. Preliminary evidence from high density genomic testing of dingoes in Ngarkat Conservation park and extending into western Victoria found evidence of limited genetic variability which is a serious conservation concern. Dingoes in Ngarkat and western Victoria had extremely low genetic variability and no evidence of gene flow with other dingo populations, demonstrating their effective isolation. This evidence suggests that the Ngarkat (and western Victorian) dingo population is threatened by inbreeding and genetic isolation. Continued culling of the Ngarkat dingo population will exacerbate the low genetic variability and threatens the persistence of this population.
We are currently seeking a Research Fellow in Eco-epidemiology/Human Ecology to join our team at Flinders University.
The successful candidate will develop spatial eco-epidemiological models for the populations of Indigenous Australians exposed to novel diseases upon contact with the first European settlers in the 18th Century. The candidate will focus on:
developing code to model how various diseases spread through and modified the demography of the Indigenous population after first contact with Europeans;
contributing to the research project by working collaboratively with the research team to deliver key project milestones;
independently contributing to ethical, high-quality, and innovative research and evaluation through activities such as scholarship, publishing in recognised, high-quality journals and assisting the preparation and submission of bids for external research funding; and
supervising of Honours and postgraduate research projects.
The ideal candidate will have advanced capacity to develop eco-epidemiological models that expand on the extensive human demographic models already developed under the auspices of the Australian Research Council Centre of Excellence for Australian Biodiversity and Heritage, of which Flinders is the Modelling Node. To be successful in this role, the candidate will demonstrate experience in coding advanced spatial models including demography, epidemiology, and ecology. The successful candidate will also demonstrate:
Thanks to the collaborative and evidence-driven foresight of my colleagues at PIRSA Biosecurity and Landscape Boards, I was recently involved in more research examining the most efficient, cost-effective, and humane ways to cull feral dear in South Australia. The resulting paper is now in review in NeoBiota, but we have also posted a pre-print of the article.
Feral deer are a real problem in Australia, and South Australia is no exception. With six species of feral deer in the country already (fallowDama dama, redCervus elaphus, hogAxis porcinus, chitalA. axis), rusaC. timorensis, and sambarRusa unicolor deer), fallow deer are the most abundant and widespread. These species are responsible for severe damage to native plants, competition with native animals, economic losses to primary industries (crops, pastures, horticulture, plantations), and human safety risks from vehicle collisions. Feral deer are also reservoirs and vectors of endemic animal diseases and have the potential to transmit exotic animal diseases such as foot-and-mouth. If left uncontrolled, within 30 years the economic impacts of feral deer could reach billions of dollars annually.
Hulton Archive / Getty Images We warn readers that the content of this study is confronting and may be distressing. In April 1789, the first smallpox epidemic among Aboriginal people in the Sydney region began, just 16 months after Europeans arrived on the First Fleet. Little is known of the true scale of the epidemic,…
Just over two years ago I reported the ‘likely’ eradication of feral pigs from Australia’s third-largest (4,405 km2) island — Kangaroo Island. I indicated ‘likely’ because the program still required the proof-of-eradication phase to be completed before an official declaration could be made. Yesterday I had the immense honour to take part in the official…
Have you ever done any research that relied to any degree on Indigenous Knowledges? How did you cite those Knowledges, if at all? It’s probably time we rethink how we engage with Indigenous Knowledge systems. In a new article published in BioScience, we — a large group of Indigenous and non-Indigenous scholars in Australia —…