Just over two years ago I reported the ‘likely’ eradication of feral pigs from Australia’s third-largest (4,405 km2) island — Kangaroo Island. I indicated ‘likely’ because the program still required the proof-of-eradication phase to be completed before an official declaration could be made.
Yesterday I had the immense honour to take part in the official declaration ceremony held on Kangaroo Island near Karatta in the island’s southwest.
I can’t begin to emphasise enough how monumental this outcome is for biodiversity conservation, not just for South Australia, or even Australia, but globally.
This is without a doubt the largest island eradication of pigs anywhere in the world. Considering that most eradication (or even density-reduction) programs fail, the success is even more outstanding.
I am also taking some personal pride in the outcome because I had a small part to play. One dreary Sunday morning back in 2022, the head of PIRSA’s Invasive Species Unit Dr Brad Page rang me with an urgent request. “Corey”, he said without much preamble, “I need a model to tell me how much it would cost to get rid of pigs on Kangaroo Island, and I need it by this afternoon”.
A few weeks ago we published a paper that was in the works for a long time, so long in fact that one of my co-authors died before it was published online.
Paul Ehrlich died on 13 March 2026 at the ripe old age of 93, exactly two weeks before our article appeared online. Paul had a good innings no doubt, but I wish he had survived long enough to see what might very well be his last co-authored paper.
I first met Paul back in the mid-2000s during a trip through San Francisco. I had organised to come chat with Professor Gretchen Daily at Stanford, and Paul came along for lunch. I remember vividly how we clicked almost immediately.
We clicked so well in fact, that we wrote a book together, co-authored several high-impact papers (e.g., ‘ghastly future‘), spent a month in Bellagio as Rockefeller Foundation writing residents, participated in various public and parliamentary presentations, and generally just got on like a house on fire. Paul and his wife Ann became like family, so much so that they were de facto grandparents to my daughter who grew up with them in near-annual contact.
This post isn’t about Paul per se, but I cannot ignore the profound influence Paul had on my career, my personality, and my life view. I miss him. I am therefore dedicating this paper and post to his memory. So long, and thanks for all the fish.
Nothing like writing about human population to get the punters engaged.
We show empirically that the Earth has already exceeded its ability to support the global human population sustainably, with dire implications for increasing pressure on food security, climate stability, and human wellbeing. However, slowing population growth and raising global awareness could still offer us some hope.
Our study shows that humans have pushed well beyond the planet’s long-term carrying capacity and that continued growth under current patterns of consumption will intensify environmental and social challenges for communities worldwide.
Procreating with a relative is taboo in most human societies for many reasons, but they all stem from avoiding one thing in particular — inbreeding increases the risk of genetic disorders that can seriously compromise a child’s health, life prospects, and survival.
While we all inherit potentially harmful mutations from our parents, the effects of these mutations are often partially or completed masked if we possess two alternative variants of a gene — one from each parent. However, the children of closely related parents are more likely to inherit the same copies of harmful mutations. This is known as ‘inbreeding depression’.
But inbreeding depression can happen in any species, with the risk increasing as populations become smaller. Because many species are rapidly declining in abundance and becoming isolated from one another predominantly due to habitat destruction, invasive species, and climate change, the chances of inbreeding are also increasing.
Not only are such populations more susceptible to random disturbances, they are also victim of reduced population growth rates arising from inbreeding depression. This produces what is generally known as the ‘extinction vortex‘ — the smaller your population, the more you inbreed and produce sub-optimal offspring, leading to even more population decline and eventually extinction.
One emergency intervention that can ‘rescue’ such inbred populations from extinction (at least in the short term) is to introduce unrelated individuals from other populations in an attempt to increase genetic diversity, and therefore, the rate of population growth. While somewhat controversial because some fear introducing diseases or eroding local-area specialisation (so-called ‘outbreeding depression’), the risk-benefit ratio of this ‘genetic rescue’ is now widely considered to be worth it.
The Black Summer bushfires of 2019–2020 that razed more than half of the landscape on Kangaroo Island in South Australia left an indelible mark on the island’s unique native biodiversity, which is still struggling to recover.
Flinders Chase National Park on Kangaroo Island after the 2019-2020 Black Summer fires (credit: CJA Bradshaw)
However, one big bonus for the environment’s recovery is the likely eradication of feral pigs (Sus scrofa). Invasive feral pigs cause a wide range of environmental, economic and social damages. In Australia, feral pigs occupy about 40% of the mainland and offshore islands, with a total, yet highly uncertain, population size estimated in the millions.
Feral pigs are recognised as a key threatening process under the Environment Protection and Biodiversity Conservation Act 1999, with impacts on at least 148 nationally threatened species and eight threatened ecological communities. They are a declared invasive species and the subject to control programs in all Australian jurisdictions.
Motion sensing cameras deployed during the eradication program capture feral pigs using their snouts to search for soil-borne food. This behaviour, called rooting, creates large areas of disturbed soil, killing native vegetation and spreading invasive weeds and pathogens (credit: PIRSA).
Imagine growing up beside the eastern Mediterranean Sea 14,000 years ago. You’re an accomplished sailor of the small watercraft you and your fellow villagers make, and you live off both the sea and the land.
But times have been difficult — there just isn’t the same amount of game or fish around as when you were a child. Maybe it’s time to look elsewhere for food.
Now imagine going farther than ever before in your little boat, accompanied maybe by a few others, when suddenly you spot something on the horizon. Is that an island?
The western coast of Cyprus. CJA Bradshaw / Flinders University
When you beach your boat to have a look around, you can’t believe what you’re seeing — tiny boar-sized hippos and horse-sized elephants that look like babies to your eyes. There are so many of them, and you’re hungry after the long journey.
The diminutive beasts don’t seem to show any fear. You easily kill a few and preserve the meat as best you can for the long journey back.
When you get home, you are excited to let everyone in the village know what you’ve found. Soon enough, you organise a major expedition back to the island.
Of course, we’ll never know if this kind of scenario took place, but it’s a plausible story of how and when the first humans managed to get to Cyprus. It also illustrates how they might have quickly brought about the demise of the tiny hippopotamusPhanourios minor, as well as the dwarf elephantPalaeoloxodon cypriotes.
Human overpopulation is often depicted in the media in one of two ways: as either a catastrophic disaster or an overly-exaggerated concern. Yet the data understood by scientists and researchers is clear. So what is the actual state of our overshoot, and, despite our growing numbers, are we already seeing the signs that the sixth mass extinction is underway?
In a recent episode of The Great Simplification podcast, Nate Hagens was joined by global ecologist Corey Bradshaw to discuss his recent research on the rapid decline in biodiversity, how population and demographics will change in the coming decades, and what both of these will mean for complex global economies currently reliant on a stable environment.
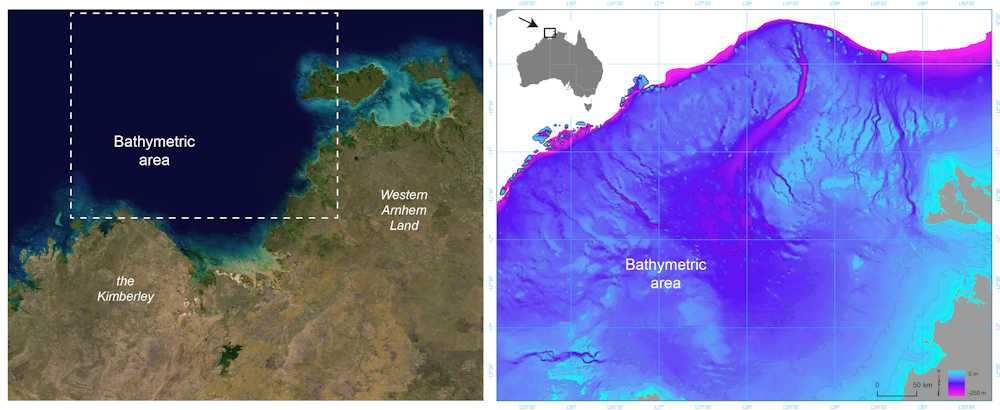
For much of the 65,000 years of Australia’s human history, the now-submerged northwest continental shelf connected the Kimberley and western Arnhem Land. This vast, habitable realm covered nearly 390,000 square kilometres, an area one-and-a-half times larger than New Zealand is today.
Left: Satellite image of the submerged northwest shelf region. Right: Drowned landscape map of the study area. US Geological Survey, Geoscience Australia
It was likely a single cultural zone, with similarities in ground stone-axe technology, styles of rock art, and languages found by archaeologists in the Kimberley and Arnhem Land.
There is plenty of archaeological evidence humans once lived on continental shelves – areas that are now submerged – all around the world. Such hard evidence has been retrieved from underwater sites in the North Sea, Baltic Sea and Mediterranean Sea, and along the coasts of North and South America, South Africa and Australia.
In a newly published study in Quaternary Science Reviews, we reveal details of the complex landscape that existed on the Northwest Shelf of Australia. It was unlike any landscape found on our continent today.
A continental split
Around 18,000 years ago, the last ice age ended. Subsequent warming caused sea levels to rise and drown huge areas of the world’s continents. This process split the supercontinent of Sahul into New Guinea and Australia, and cut Tasmania off from the mainland.
Unlike in the rest of the world, the now-drowned continental shelves of Australia were thought to be environmentally unproductive and little used by First Nations peoples.
But mounting archaeological evidence shows this assumption is incorrect. Many large islands off Australia’s coast – islands that once formed part of the continental shelves – show signs of occupation before sea levels rose.
Stone tools have also recently been found on the sea floor off the coast of the Pilbara region of Western Australia.
Have you ever watched a nature documentary and marvelled at the intricate dance of life unfolding on screen? From the smallest insect to the largest predator, every creature plays a role in the grand performance of our planet’s biosphere. But what happens when one of these performers disappears?
In this post, we delve into our recent article Estimating co-extinction risks in terrestrial ecosystems just published in Global Change Biology, in which we discuss the cascading effects of species loss and the risks of ‘co-extinction’.
But what does ‘co-extinction’ really mean?
Imagine an ecosystem as a giant web of interconnected species. Each thread represents a relationship between two species — for example, a bird that eats a certain type of insect, or a plant that relies on a specific species of bee for pollination. Now, what happens if one of these species in the pair disappears? The thread breaks and the remaining species loses an interaction. This could potentially lead to its co-extinction, which is essentially the domino effect of multiple species losses in an ecosystem.
A famous example of this effect can be seen with the invasion of the cane toad (Rhinella marina) across mainland Australia, which have caused trophic cascades and species compositional changes in these communities.
The direct extinction of one species, caused by effects such as global warming for example, has the potential to cause other species also to become extinct indirectly.
We are currently seeking a Research Fellow in Eco-epidemiology/Human Ecology to join our team at Flinders University.
The successful candidate will develop spatial eco-epidemiological models for the populations of Indigenous Australians exposed to novel diseases upon contact with the first European settlers in the 18th Century. The candidate will focus on:
developing code to model how various diseases spread through and modified the demography of the Indigenous population after first contact with Europeans;
contributing to the research project by working collaboratively with the research team to deliver key project milestones;
independently contributing to ethical, high-quality, and innovative research and evaluation through activities such as scholarship, publishing in recognised, high-quality journals and assisting the preparation and submission of bids for external research funding; and
supervising of Honours and postgraduate research projects.
The ideal candidate will have advanced capacity to develop eco-epidemiological models that expand on the extensive human demographic models already developed under the auspices of the Australian Research Council Centre of Excellence for Australian Biodiversity and Heritage, of which Flinders is the Modelling Node. To be successful in this role, the candidate will demonstrate experience in coding advanced spatial models including demography, epidemiology, and ecology. The successful candidate will also demonstrate:
Wildfires transform forests into mosaics of vegetation. What, where, and which plants thrive depends on when and how severely a fire affects different areas of a forest. Such heterogeneity in the landscape is essential for animal species that benefit from fire like woodpeckers.
The black-backed woodpecker (Picoides arcticus) lives in the coniferous forests of North America’s boreal-Mediterranean region. Thanks to a powerful and sharp bill, this bird can excavate nests inside the trunks of (mainly dead) trees, and those cavities will be re-used later by many species of birds, mammals, and invertebrates in fire-prone landscapes (22). The images show a male with the characteristic black plumage of his back that serves as camouflage against the dark bark of a dead tree three years after a wildfire in Montana (USA). Being omnivores, the diet of this bird largely relies on the larvae of woodboring coleoptera like jewell and longhorn beetles. These insects are abundant post-fire, the champion being the fire beetle (Melanophila spp.). The thorax of fire beetles is equipped with infrared-light receptors that can detect a wildfire from tens of kilometres away (23). These fascinating little beasts are the first to arrive at a burned forest and, of course, woodpeckers follow soon after. The preference of the blackbacked woodpecker for burned forests and their cryptic feathers and pyrophilic diet reflect a long evolutionary history in response to fires. Courtesy of Richard Hutto.
Anyone raised in rural areas will have vivid recollections of wildfires: the thick, ashy smell, the overcast sky on a sunny day, and the purring of aerial firefighters dropping water from their hanging tanks. The reality is that wildfires are natural events that shape biodiversity and ecosystem function (1) — to the extent that fire is intimately linked to the appearance and evolution of terrestrial plants (2). Since the Palaeolithic, our own species has used fire at will, to cook, hunt, melt metals, open cropland or paths, or tell stories in front of a hearth (3).
Where there are regular wildfires (fire-prone ecosystems), different areas of the landscape burn in different seasons and years under different weather patterns. Therefore, each region has a unique fire biography in terms of how frequently, how much, and how long ago wildfires occurred. All those factors interact will one another and with topography.
Flooding in the Murray-Darling Basin is creating ideal breeding conditions for many native species that have evolved to take advantage of temporary flood conditions. Led by PhD candidate Rupert Mathwin, our team developed virtual models of the Murray River to reveal a crucial link between natural flooding and the extinction risk of endangered southern bell frogs (Litoria raniformis; also known as growling grass frogs).
Southern bell frogs are one of Australia’s 100 Priority Threatened Species. This endangered frog breeds during spring and summer when water levels increase in their wetlands. However, the natural flooding patterns in Australia’s largest river system have been negatively impacted by expansive river regulation that some years, sees up to 60% of river water extracted for human use.
Our latest paper describes how we built computer simulations of Murray-Darling Basin wetlands filled with simulated southern bell frogs. By changing the simulation from natural to regulated conditions, we showed that modern conditions dramatically increase the extinction risk of these beloved frogs.
The data clearly indicate that successive dry years raise the probability of local extinction, and these effects are strongest in smaller wetlands. Larger wetlands and those with more frequent inundation are less prone to these effects, although they are not immune to them entirely. The models present a warning — we have greatly modified the way the river behaves, and the modern river cannot support the long-term survival of southern bell frogs.’
Last week, researchers at the University of Melbourne announced that thylacines or Tasmanian tigers, the Australian marsupial predators extinct since the 1930s, could one day be ushered back to life.
The thylacine (Thylacinus cynocephalus), also known as the ‘Tasmanian tiger’ (it was neither Tasmanian, because it was once common in mainland Australia, nor was it related to the tiger), went extinct in Tasmania in the 1930s from persecution by farmers and habitat loss. Art by Eleanor (Nellie) Pease, University of Queensland. Centre of Excellence for Australian Biodiversity and Heritage
Advances in mapping the genome of the thylacine and its living relative the numbat have made the prospect of re-animating the species seem real. As an ecologist, I would personally relish the opportunity to see a living specimen.
The announcement led to some overhyped headlines about the imminent resurrection of the species. But the idea of “de-extinction” faces a variety of technical, ethical and ecological challenges. Critics (like myself) argue it diverts attention and resources from the urgent and achievable task of preventing still-living species from becoming extinct.
The rebirth of the bucardo
The idea of de-extinction goes back at least to the the creation of the San Diego Frozen Zoo in the early 1970s. This project aimed to freeze blood, DNA, tissue, cells, eggs and sperm from exotic and endangered species in the hope of one day recreating them.
The notion gained broad public attention with the first of the Jurassic Park films in 1993. The famous cloning of Dolly the sheep reported in 1996 created a sense that the necessary know-how wasn’t too far off.
The next technological leap came in 2008, with the cloning of a dead mouse that had been frozen at –20℃ for 16 years. If frozen individuals could be cloned, re-animation of a whole species seemed possible.
After this achievement, de-extinction began to look like a potential way to tackle the modern global extinction crisis.
Back in June of this year I wrote (whinged) about the disappointment of writing a lot of ecological models that were rarely used to assist real-world wildlife management. However, I did hint that another model I wrote had assistance one government agency with pig management on Kangaroo Island.
Modelling by the Flinders UniversityGlobal Ecology Laboratory shows the likelihood and feasibility of feral pig eradication under different funding and eradication scenarios. With enough funding, feral pigs could be eradicated from Kangaroo Island in 2 years.
This basically means that because of the model, PIRSA was successful in obtaining enough funding to pretty much ensure that the eradication of feral pigs from Kangaroo Island will be feasible!
Why is this important to get rid of feral pigs? They are a major pest on the Island, causing severe economic and environmental impacts both to farms and native ecosystems. On the agricultural side of things, they prey on newborn lambs, eat crops, and compete with livestock for pasture. Feral pigs damage natural habitats by up-rooting vegetation and fouling waterholes. They can also spread weeds and damage infrastructure, as well as act as hosts of parasites and diseases (e.g., leptospirosis, tuberculosis, foot-and-mouth disease) that pose serious threats to industry, wildlife, and even humans.
After the rather astounding response to our Ghastly Future paper published in January this year (> 443,000 views and counting; 61 citations and counting), we received a Commentary that was rather critical of our article.
A Malthusian slur
We have finally published a Response to the Commentary, which is now available online (accepted version) in Frontiers in Conservation Science. Given that it is published under a Creative Commons Attribution License (CC BY), I can repost the Response here:
In their comment on our paper Underestimating the challenges of avoiding a ghastly future, Bluwstein et al.2 attempt to contravene our exposé of the enormous challenges facing the entire human population from a rapidly degrading global environment. While we broadly agree with the need for multi-disciplinary solutions, and we worry deeply about the inequality of those who pay the costs of biodiversity loss and ecological collapse, we feel obligated to correct misconceptions and incorrect statements that Bluwstein et al.2 made about our original article.
After incorrectly assuming that our message implied the existence of “one science” and a “united scientific community”, the final paragraph of their comment contradicts their own charge by calling for the scientific community to “… stand in solidarity”. Of course, there is no “one science” — we never made such a claim. Science is by its nature necessarily untidy because it is a bottom-up process driven by different individuals, cultures, perspectives, and goals. But it is solid at the core. Scientific confluence is reached by curiosity, rigorous testing of assumptions, and search for contradictions, leading to many — sometimes counter-intuitive or even conflicting — insights about how the world works. There is no one body of scientific knowledge, even though there is good chance that disagreements are eventually resolved by updated, better evidence, although perhaps too slowly. That was, in fact, a main message of our original article — that obligatory specialisation of disparate scientific fields, embedded within a highly unequal and complex socio-cultural-economic framework, reduces the capacity of society to appreciate, measure, and potentially counter the complexity of its interacting existential challenges. We agree that scientists play a role in political struggles, but we never claimed, as Bluwstein et al.2 contended, that such struggles can be “… reduced to science-led processes of positive change”. Indeed, this is exactly the reason our paper emphasized the political impotence surrounding the required responses. We obviously recognize the essential role social scientists play in creating solutions to avoid a ghastly future. Science can only provide the best available evidence that individuals and policymakers can elect to use to inform their decisions.
We certainly recognise that there is no single policy or polity capable of addressing compounding and mounting problems, and we agree that that there is no “universal understanding of the intertwined socio-ecological challenges we face”. Bluwstein et al.2 claimed that we had suggested scientific messaging alone can “… adequately communicate to the public how socio-ecological crises should be addressed”. We did not state or imply such ideas of unilateral scientific power anywhere in our article. Indeed, the point of framing our message as pertaining to a complex adaptive system means that we cannot, and should not, work towards a single goal. Instead, humanity will be more successful tackling challenges simultaneously and from multiple perspectives, by exploiting manifold institutions, technologies, approaches, and governances to match the complexity of the predicament we are attempting to resolve.
I’m pleased to announce the publication of a paper led by Kathryn Venning (KV) that was derived from her Honours work in the lab. Although she’s well into her PhD on an entirely different topic, I’m overjoyed that she persevered and saw this work to publication.
Feral cats occupy every habitat in the country, from the high tropics to the deserts, and from the mountains to the sea. They adapt to the cold just as easily as they adapt to the extreme heat, and they can eat just about anything that moves, from invertebrates to the carcases of much larger animals that they scavenge.
Cats are Australia’s bane, but you can’t help but be at least a little impressed with their resilience.
Still, we have to try our best to get rid of them where we can, or at least reduce their densities to the point where their ecological damage is limited.
Typically, the only efficient and cost-effective way to do that is via lethal control, but by using various means. These can include direct shooting, trapping, aerial poison-baiting, and a new ‘smart’ method of targeted poison delivery via a prototype device known as a Felixer™️. The latter are particularly useful for passive control in areas where ground-shooting access is difficult.
A live Felixer™️ deployed on Kangaroo Island (photo: CJA Bradshaw 2020)
A few years back the federal government committed what might seem like a sizeable amount of money to ‘eradicate’ cats from Australia. Yeah, good luck with that, although the money has been allocated to several places where cat reduction and perhaps even eradication is feasible. Namely, on islands.
We know it is more than 60,000 years since the first people entered the continent of Sahul — the giant landmass that connected New Guinea, Australia and Tasmania when sea levels were lower than today.
But where the earliest people moved across the landscape, how fast they moved, and how many were involved, have been shrouded in mystery.
Our latest research, published today shows the establishment of populations in every part of this giant continent could have occurred in as little as 5,000 years. And the entire population of Sahul could have been as high as 6.4 million people.
This translates to more than 3 million people in the area that is now modern-day Australia, far more than any previous estimate.
The first people could have entered through what is now western New Guinea or from the now-submerged Sahul Shelf off the modern-day Kimberley (or both).
But whichever the route, entire communities of people arrived, adapted to and established deep cultural connections with Country over 11 million square kilometres of land, from northwestern Sahul to Tasmania.
Map of what Australia looked like for most of the human history of the continent when sea levels were lower than today. Author provided
This equals a rate of population establishment of about 1km per year (based on a maximum straight-line distance of about 5,000km from the introduction point to the farthest point).
That’s doubly impressive when you consider the harshness of the Australian landscape in which people both survived and thrived.
For many years I’ve been interested in modelling the extinction dynamics of megafauna. Apart from co-authoring a few demographically simplified (or largely demographically free) models about how megafauna species could have gone extinct, I have never really tried to capture the full nuances of long-extinct species within a fully structured demographic framework.
That is, until now.
But how do you get the life-history data of an extinct animal that was never directly measured. Surely, things like survival, reproductive output, longevity and even environmental carrying capacity are impossible to discern, and aren’t these necessary for a stage-structured demographic model?
The answer to the first part of that question “it’s possible”, and to the second, it’s “yes”. The most important bit of information we palaeo modellers need to construct something that’s ecologically plausible for an extinct species is an estimate of body mass. Thankfully, palaeontologists are very good at estimating the mass of the things they dig up (with the associated caveats, of course). From such estimates, we can reconstruct everything from equilibrium densities, maximum rate of population growth, age at first breeding, and longevity.
But it’s more complicated than that, of course. In Australia anyway, we’re largely dealing with marsupials (and some monotremes), and they have a rather different life-history mode than most placentals. We therefore have to ‘correct’ the life-history estimates derived from living placental species. Thankfully, evolutionary biologists and ecologists have ways to do that too.
The Pleistocene kangaroo Procoptodon goliah, the largest and most heavily built of the short-faced kangaroos, was the largest and most heavily built kangaroo known. It had an unusually short, flat face and forwardly directed eyes, with a single large toe on each foot (reduced from the more normal count of four). Each forelimb had two long, clawed fingers that would have been used to bring leafy branches within reach.
So with a battery of ecological, demographic, and evolutionary tools, we can now create reasonable stochastic-demographic models for long-gone species, like wombat-like creatures as big as cars, birds more than two metres tall, and lizards more than seven metres long that once roamed the Australian continent.
Ancient clues, in the shape of fossils and archaeological evidence of varying quality scattered across Australia, have formed the basis of several hypotheses about the fate of megafauna that vanished during a peak about 42,000 years ago from the ancient continent of Sahul, comprising mainland Australia, Tasmania, New Guinea and neighbouring islands.
There is a growing consensus that multiple factors were at play, including climate change, the impact of people on the environment, and access to freshwater sources.
Just published in the open-access journal eLife, our latest CABAH paper applies these approaches to assess how susceptible different species were to extinction – and what it means for the survival of species today.
Using various characteristics such as body size, weight, lifespan, survival rate, and fertility, we (Chris Johnson, John Llewelyn, Vera Weisbecker, Giovanni Strona, Frédérik Saltré & me) created population simulation models to predict the likelihood of these species surviving under different types of environmental disturbance.
We compared the results to what we know about the timing of extinction for different megafauna species derived from dated fossil records. We expected to confirm that the most extinction-prone species were the first species to go extinct – but that wasn’t necessarily the case.
While we did find that slower-growing species with lower fertility, like the rhino-sized wombat relative Diprotodon, were generally more susceptible to extinction than more-fecund species like the marsupial ‘tiger’ thylacine, the relative susceptibility rank across species did not match the timing of their extinctions recorded in the fossil record.
Indeed, we found no clear relationship between a species’ inherent vulnerability to extinction — such as being slower and heavier and/or slower to reproduce — and the timing of its extinction in the fossil record.
In fact, we found that most of the living species used for comparison — such as short-beaked echidnas, emus, brush turkeys, and common wombats — were more susceptible on average than their now-extinct counterparts.
In some African countries, lion trophy hunting is legal. Riaan van den Berg
In sub-Saharan Africa, almost 1,400,000 km² of land spread across many countries — from Kenya to South Africa — is dedicated to “trophy” (recreational) hunting. This type of hunting can occur on communal, private, and state lands.
The hunters – mainly foreign “tourists” from North America and Europe – target a wide variety of species, including lions, leopards, antelopes, buffalo, elephants, zebras, hippopotamus and giraffes.
Debates centred on the role of recreational hunting in supporting nature conservation and local people’s livelihoods are among the most polarising in conservation today.
On one hand, people argue that recreational hunting generates funding that can support livelihoods and nature conservation. It’s estimated to generate US$200 million annually in sub-Saharan Africa, although others dispute the magnitude of this contribution.
On the other hand, hunting is heavily criticised on ethical and moral grounds and as a potential threat to some species.
Evidence for taking a particular side in the debate is still unfortunately thin. In our recently published research, we reviewed the large body of scientific literature on recreational hunting from around the world, which meant we read and analysed more than 1000 peer-reviewed papers.
My father was a hunter, and by proxy so was I when I was a lad. I wasn’t really a ‘good’ hunter in the sense that I rarely bagged my quarry, but during my childhood not only did I fail to question the morality of recreational hunting, I really thought that in fact it was by and large an important cultural endeavour.
It’s interesting how conditioned we become as children, for I couldn’t possibly conceive of hunting a wild, indigenous species for my own personal satisfaction now. I find the process not only morally and ethically reprehensible, I also think that most species don’t need the extra stress in an already environmentally stressed world.
I admit that I do shoot invasive European rabbits and foxes on my small farm from time to time — to reduce the grazing and browsing pressure on my trees from the former, and the predation pressure on the chooks from the latter. Of course, we eat the rabbits, but I tend just to bury the foxes. My dual perspective on the general issue of hunting in a way mirrors the two sides of the recreational hunting issue we report in our latest paper.
Wild boar (Sus scrofus). Photo: Valentin Panzirsch, CC BY-SA 3.0 AT, via Wikimedia Commons
I want to be clear here that our paper focuses exclusively on recreational hunting, and especially the hunting of charismatic species for their trophies. The activity is more than just a little controversial, for it raises many ethical and moral concerns at the very least. Yet, recreational hunting is frequently suggested as a way to conserve nature and support local people’s livelihoods.
However, this time I’ve strayed from my recent bibliometric musings and developed something that’s more compatible with the core of my main research and interests.
Over the years I’ve taught many students the basics of population modelling, with the cohort-based approaches dominating the curriculum. Of these, the simpler ‘Leslie’ (age-classified) matrix models are both the easiest to understand and for which data can often be obtained without too many dramas.
But unless you’re willing to sit down and learn the code, they can be daunting to the novice.
Sure, there are plenty of software alternatives out there, such as Bob Lacy‘s Vortex (a free individual-based model available for PCs only), Resit Akçakaya & co’s RAMAS Metapop ($; PC only), Stéphane Legendre‘s Unified Life Models (ULM; open-source; all platforms), and Charles Todd‘s Essential (open-source; PC only) to name a few. If you’re already an avid R user and already into population modelling, you might be familiar with the population-modelling packages popdemo, OptiPopd, or sPop. I’m sure there are still other good resources out there of which I’m not aware.
But, even to install the relevant software or invoke particular packages in R takes a bit of time and learning. It’s probably safe to assume that many people find the prospect daunting.
It’s for this reason that I turned my newly acquired R Shiny skills to matrix population models so that even complete coding novices can run their own stochastic population models.
Get Conservation Bytes by Email Enter your email address to receive new posts on conservation, biodiversity, and environmental science directly in your inbox. No need to visit the website regularly, and you can unsubscribe at any time.
Hulton Archive / Getty Images We warn readers that the content of this study is confronting and may be distressing. In April 1789, the first smallpox epidemic among Aboriginal people in the Sydney region began, just 16 months after Europeans arrived on the First Fleet. Little is known of the true scale of the epidemic,…
Just over two years ago I reported the ‘likely’ eradication of feral pigs from Australia’s third-largest (4,405 km2) island — Kangaroo Island. I indicated ‘likely’ because the program still required the proof-of-eradication phase to be completed before an official declaration could be made. Yesterday I had the immense honour to take part in the official…
Have you ever done any research that relied to any degree on Indigenous Knowledges? How did you cite those Knowledges, if at all? It’s probably time we rethink how we engage with Indigenous Knowledge systems. In a new article published in BioScience, we — a large group of Indigenous and non-Indigenous scholars in Australia —…



























The very worn slur of “neo-Malthusian”
7 09 2021After the rather astounding response to our Ghastly Future paper published in January this year (> 443,000 views and counting; 61 citations and counting), we received a Commentary that was rather critical of our article.
We have finally published a Response to the Commentary, which is now available online (accepted version) in Frontiers in Conservation Science. Given that it is published under a Creative Commons Attribution License (CC BY), I can repost the Response here:
In their comment on our paper Underestimating the challenges of avoiding a ghastly future, Bluwstein et al.2 attempt to contravene our exposé of the enormous challenges facing the entire human population from a rapidly degrading global environment. While we broadly agree with the need for multi-disciplinary solutions, and we worry deeply about the inequality of those who pay the costs of biodiversity loss and ecological collapse, we feel obligated to correct misconceptions and incorrect statements that Bluwstein et al.2 made about our original article.
After incorrectly assuming that our message implied the existence of “one science” and a “united scientific community”, the final paragraph of their comment contradicts their own charge by calling for the scientific community to “… stand in solidarity”. Of course, there is no “one science” — we never made such a claim. Science is by its nature necessarily untidy because it is a bottom-up process driven by different individuals, cultures, perspectives, and goals. But it is solid at the core. Scientific confluence is reached by curiosity, rigorous testing of assumptions, and search for contradictions, leading to many — sometimes counter-intuitive or even conflicting — insights about how the world works. There is no one body of scientific knowledge, even though there is good chance that disagreements are eventually resolved by updated, better evidence, although perhaps too slowly. That was, in fact, a main message of our original article — that obligatory specialisation of disparate scientific fields, embedded within a highly unequal and complex socio-cultural-economic framework, reduces the capacity of society to appreciate, measure, and potentially counter the complexity of its interacting existential challenges. We agree that scientists play a role in political struggles, but we never claimed, as Bluwstein et al.2 contended, that such struggles can be “… reduced to science-led processes of positive change”. Indeed, this is exactly the reason our paper emphasized the political impotence surrounding the required responses. We obviously recognize the essential role social scientists play in creating solutions to avoid a ghastly future. Science can only provide the best available evidence that individuals and policymakers can elect to use to inform their decisions.
We certainly recognise that there is no single policy or polity capable of addressing compounding and mounting problems, and we agree that that there is no “universal understanding of the intertwined socio-ecological challenges we face”. Bluwstein et al.2 claimed that we had suggested scientific messaging alone can “… adequately communicate to the public how socio-ecological crises should be addressed”. We did not state or imply such ideas of unilateral scientific power anywhere in our article. Indeed, the point of framing our message as pertaining to a complex adaptive system means that we cannot, and should not, work towards a single goal. Instead, humanity will be more successful tackling challenges simultaneously and from multiple perspectives, by exploiting manifold institutions, technologies, approaches, and governances to match the complexity of the predicament we are attempting to resolve.
Read the rest of this entry »Share:
Comments : Leave a Comment »
Tags: commentary, complex adaptive system, consumption, critique, human population, Malthusian, neo-Malthusian, over-population, overshoot, Population
Categories : agriculture, anthropocene, biodiversity, climate change, demography, economics, education, Endarkenment, environmental economics, environmental policy, extinction, food, governance, human overpopulation, poverty, science, societies, sustainability