Procreating with a relative is taboo in most human societies for many reasons, but they all stem from avoiding one thing in particular — inbreeding increases the risk of genetic disorders that can seriously compromise a child’s health, life prospects, and survival.
While we all inherit potentially harmful mutations from our parents, the effects of these mutations are often partially or completed masked if we possess two alternative variants of a gene — one from each parent. However, the children of closely related parents are more likely to inherit the same copies of harmful mutations. This is known as ‘inbreeding depression’.
But inbreeding depression can happen in any species, with the risk increasing as populations become smaller. Because many species are rapidly declining in abundance and becoming isolated from one another predominantly due to habitat destruction, invasive species, and climate change, the chances of inbreeding are also increasing.
Not only are such populations more susceptible to random disturbances, they are also victim of reduced population growth rates arising from inbreeding depression. This produces what is generally known as the ‘extinction vortex‘ — the smaller your population, the more you inbreed and produce sub-optimal offspring, leading to even more population decline and eventually extinction.
One emergency intervention that can ‘rescue’ such inbred populations from extinction (at least in the short term) is to introduce unrelated individuals from other populations in an attempt to increase genetic diversity, and therefore, the rate of population growth. While somewhat controversial because some fear introducing diseases or eroding local-area specialisation (so-called ‘outbreeding depression’), the risk-benefit ratio of this ‘genetic rescue’ is now widely considered to be worth it.
Nearly a decade ago (my how time flies*), I wrote a post about the guaranteed failure of government policies purporting no-extinction targets within their environmental plans. I was referring to the State of South Australia’s (then) official policy of no future extinctions.
In summary, zero- (or no-) extinction targets at best demonstrate a deep naïvety of how ecology works, and at worst, waste a lot of resources on interventions doomed to fail.
4. Climate change will also guarantee additional (perhaps even most) future extinctions irrespective of Australian policies.
I argued that no-extinction policies are therefore disingenuous to the public in the extreme because they sets false expectations, engender disillusionment after inevitable failure, and ignores the concept of triage — putting our environment-restoration resources toward the species/systems with the best chance of surviving (uniqueness notwithstanding).
When snorkelling in a reef, it’s natural to think of coral colonies as a colourful scenography where fish act in a play. But what would happen to the fish if the stage went suddenly empty, as in Peter Brook’s 1971 Midsummer Night’s Dream? Would the fish still be there acting their roles without a backdrop?
This question is not novel in coral-reef science. Ecologists have often compared reef fish diversity and biomass in selected localities before and after severe events of coral mortality. Even a temporary disappearance of corals might have substantial effects on fish communities, sometimes resulting in a local disappearance of more than half of local fish species.
Considering the multiple, complex ways fish interact with — and depend on — corals, this might appear as an obvious outcome. Still, such complexity of interactions makes it difficult to predict how the loss of corals might affect fish diversity in specific contexts, let alone at the global scale.
Focusing on species-specific fish-coral associations reveals an inconsistent picture with local-scale empirical observations. When looking at the fraction of local fish diversity that strictly depends on corals for food and other more generic habitat requirements (such as shelter and reproduction), the global picture suggests that most fish diversity in reef locality might persist in the absence of corals.
The mismatch between this result and the empirical evidence of a stronger coral dependence suggests the existence of many hidden ecological paths connecting fish to corals, and that those paths might entrap many fish species for which the association to corals is not apparent.
For many years I’ve been interested in modelling the extinction dynamics of megafauna. Apart from co-authoring a few demographically simplified (or largely demographically free) models about how megafauna species could have gone extinct, I have never really tried to capture the full nuances of long-extinct species within a fully structured demographic framework.
That is, until now.
But how do you get the life-history data of an extinct animal that was never directly measured. Surely, things like survival, reproductive output, longevity and even environmental carrying capacity are impossible to discern, and aren’t these necessary for a stage-structured demographic model?
The answer to the first part of that question “it’s possible”, and to the second, it’s “yes”. The most important bit of information we palaeo modellers need to construct something that’s ecologically plausible for an extinct species is an estimate of body mass. Thankfully, palaeontologists are very good at estimating the mass of the things they dig up (with the associated caveats, of course). From such estimates, we can reconstruct everything from equilibrium densities, maximum rate of population growth, age at first breeding, and longevity.
But it’s more complicated than that, of course. In Australia anyway, we’re largely dealing with marsupials (and some monotremes), and they have a rather different life-history mode than most placentals. We therefore have to ‘correct’ the life-history estimates derived from living placental species. Thankfully, evolutionary biologists and ecologists have ways to do that too.
The Pleistocene kangaroo Procoptodon goliah, the largest and most heavily built of the short-faced kangaroos, was the largest and most heavily built kangaroo known. It had an unusually short, flat face and forwardly directed eyes, with a single large toe on each foot (reduced from the more normal count of four). Each forelimb had two long, clawed fingers that would have been used to bring leafy branches within reach.
So with a battery of ecological, demographic, and evolutionary tools, we can now create reasonable stochastic-demographic models for long-gone species, like wombat-like creatures as big as cars, birds more than two metres tall, and lizards more than seven metres long that once roamed the Australian continent.
Ancient clues, in the shape of fossils and archaeological evidence of varying quality scattered across Australia, have formed the basis of several hypotheses about the fate of megafauna that vanished during a peak about 42,000 years ago from the ancient continent of Sahul, comprising mainland Australia, Tasmania, New Guinea and neighbouring islands.
There is a growing consensus that multiple factors were at play, including climate change, the impact of people on the environment, and access to freshwater sources.
Just published in the open-access journal eLife, our latest CABAH paper applies these approaches to assess how susceptible different species were to extinction – and what it means for the survival of species today.
Using various characteristics such as body size, weight, lifespan, survival rate, and fertility, we (Chris Johnson, John Llewelyn, Vera Weisbecker, Giovanni Strona, Frédérik Saltré & me) created population simulation models to predict the likelihood of these species surviving under different types of environmental disturbance.
We compared the results to what we know about the timing of extinction for different megafauna species derived from dated fossil records. We expected to confirm that the most extinction-prone species were the first species to go extinct – but that wasn’t necessarily the case.
While we did find that slower-growing species with lower fertility, like the rhino-sized wombat relative Diprotodon, were generally more susceptible to extinction than more-fecund species like the marsupial ‘tiger’ thylacine, the relative susceptibility rank across species did not match the timing of their extinctions recorded in the fossil record.
Indeed, we found no clear relationship between a species’ inherent vulnerability to extinction — such as being slower and heavier and/or slower to reproduce — and the timing of its extinction in the fossil record.
In fact, we found that most of the living species used for comparison — such as short-beaked echidnas, emus, brush turkeys, and common wombats — were more susceptible on average than their now-extinct counterparts.
Anyone with even a passing interest in the global environment knows all is not well. But just how bad is the situation? Our new paper shows the outlook for life on Earth is more dire than is generally understood.
The research published today reviews more than 150 studies to produce a stark summary of the state of the natural world. We outline the likely future trends in biodiversity decline, mass extinction, climate disruption and planetary toxification. We clarify the gravity of the human predicament and provide a timely snapshot of the crises that must be addressed now.
The problems, all tied to human consumption and population growth, will almost certainly worsen over coming decades. The damage will be felt for centuries and threatens the survival of all species, including our own.
Our paper was authored by 17 leading scientists, including those from Flinders University, Stanford University and the University of California, Los Angeles. Our message might not be popular, and indeed is frightening. But scientists must be candid and accurate if humanity is to understand the enormity of the challenges we face.
Humanity must come to terms with the future we and future generations face. Shutterstock
Getting to grips with the problem
First, we reviewed the extent to which experts grasp the scale of the threats to the biosphere and its lifeforms, including humanity. Alarmingly, the research shows future environmental conditions will be far more dangerous than experts currently believe. Read the rest of this entry »
Around a fortnight ago I wrote a hastily penned post about the precarious state of biodiversity — it turned out to be one of the most-read posts in ConservationBytes‘ history (nearly 22,000 views in less than two weeks).
Now, let’s examine whether this dreadful history is likely to get any better any time soon.
Even if extinction rates decline substantially over the next century, I argue that we are committed to an intensifying biodiversity extinction crisis. The aggregate footprint from the growing human population notwithstanding, we can expect decades, if not centuries, of continued extinctions from lag effects alone (extinction debts arising from previous environmental damage engendering extinctions in the future)1.
Global vegetation cover and production are also likely to decline even in the absence of continued habitat clearing — the potential benefit of higher CO2 concentrations for plant photosynthesis is more than offset by lower availability of water in the soil, heat stress, and the frequency of disturbances such as droughts2. Higher frequencies and intensities of disturbance events like catastrophic bushfire will also exacerbate extinction rates3.
However, perhaps the least-appreciated element of potential extinctions arising from climate change is that they are vastly underestimated when only considering a species’ thermal tolerance4. In fact, climate disruption-driven extinction rates could be up to ten times higher than currently predicted4 when extinction cascades are taken into account5. Read the rest of this entry »
As I’ve done for the last six years, I am publishing a retrospective list of the ‘top’ 20 influential papers of 2019 as assessed by experts in F1000 Prime (in no particular order). See previous years’ lists here: 2018, 2017, 2016, 2015, 2014, and 2013.
For more than 3.5 billion years, living organisms have thrived, multiplied and diversified to occupy every ecosystem on Earth. The flip side to this explosion of new species is that species extinctions have also always been part of the evolutionary life cycle.
But these two processes are not always in step. When the loss of species rapidly outpaces the formation of new species, this balance can be tipped enough to elicit what are known as “mass extinction” events.
A mass extinction is usually defined as a loss of about three quarters of all species in existence across the entire Earth over a “short” geological period of time. Given the vast amount of time since life first evolved on the planet, “short” is defined as anything less than 2.8 million years.
Since at least the Cambrian periodthat began around 540 million years ago when the diversity of life first exploded into a vast array of forms, only five extinction events have definitively met these mass-extinction criteria.
These so-called “Big Five” have become part of the scientific benchmark to determine whether human beings have today created the conditions for a sixth mass extinction.
An ammonite fossil found on the Jurassic Coast in Devon. The fossil record can help us estimate prehistoric extinction rates.Corey Bradshaw, Author provided
Here’s a presentation I gave earlier in the year for the Flinders University BRAVE Research and Innovation series:
There is No Plan(et) B — What you can do about Earth’s extinction emergency
Earth is currently experiencing a mass extinction brought about by, … well, … us. Species are being lost at a rate similar to when the dinosaurs disappeared. But this time, it’s not due to a massive asteroid hitting the Earth; species are being removed from the planet now because of human consumption of natural resources. Is a societal collapse imminent, and do we need to prepare for a post-collapse society rather than attempt to avoid one? Or, can we limit the severity and onset of a collapse by introducing a few changes such as removing political donations, becoming vegetarians, or by reducing the number of children one has?
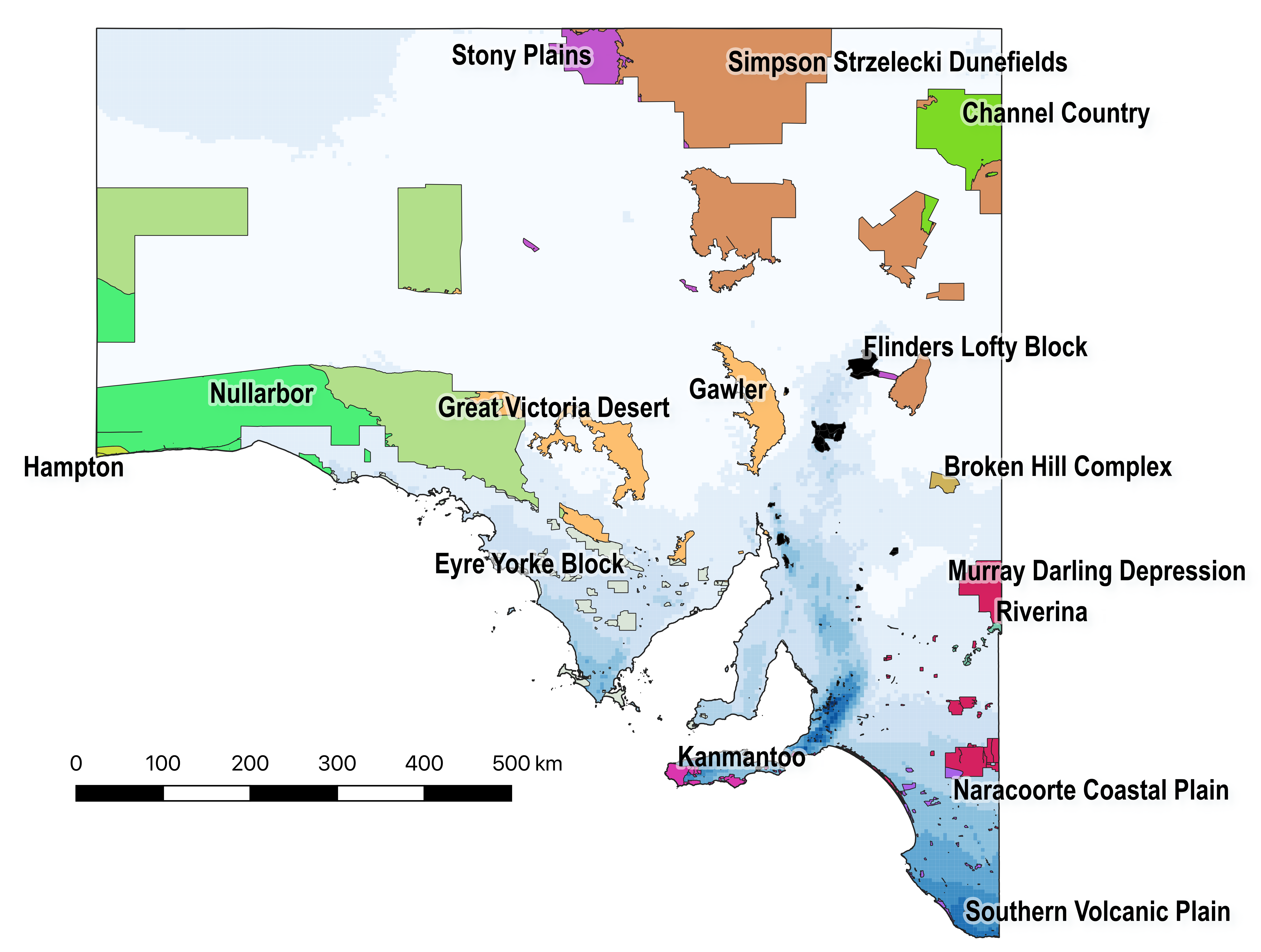
Figure 2 (from the article). Overlaying the South Australia’s Protected Areas boundary data with the Interim Biogeographic Regionalisation for Australia layer indicates that 73.2% of the total protected area (excluding Indigenous Protected Areas) in South Australia lies in the arid biogeographic regions of Great Victoria Desert (21.1%), Channel Country (15.2%), Simpson Strzelecki Dunefields (14.0%), Nullarbor (9.8%), Stony Plains (6.6%), Gawler (6.0%), and Hampton (0.5%). The total biogeographic-region area covered by the remaining Conservation Reserves amounts to 26.2%. Background blue shading indicates relative average annual rainfall.
If you read CB.com regularly, you’ll know that late last year I blogged about the South Australia 2108 State of the Environment Report for which I was commissioned to write an ‘overview‘ of the State’s terrestrial biodiversity.
At the time I whinged that not many people seemed to take notice (something I should be used to by now in the age of extremism and not giving a tinker’s about the future health of the planet — but I digress), but it seems that quietly, quietly, at least people with some policy influence here are starting to listen.
Not satisfied with merely having my report sit on the virtual shelves at the SA Environment Protection Authority, I decided that I should probably flesh out the report and turn it into a full, peer-reviewed article.
The paper is chock-a-block with all the same sorts of points I covered last year, but there’s a lot more, and it’s also a lot better referenced and logically sequenced.
For the last five years I’ve published a retrospective list of the ‘top’ 20 influential papers of the year as assessed by experts in F1000 Prime — so, I’m doing so again for 2018 (interesting side note: six of the twenty papers highlighted here for 2018 appear in Science magazine). See previous years’ posts here: 2017, 2016, 2015, 2014, and 2013.
Just under two weeks ago, Giovanni Strona and I published a paper in Scientific Reports on measuring the co-extinction effect from climate change. What we found even made me — an acknowledged pessimist — stumble in shock and incredulity.
But a bit of back story is necessary before I launch into describing what we discovered.
Last year, some Oxbridge astrophysicists (David Sloan and colleagues) published a rather sensational paper in Scientific Reports claiming that life on Earth would likely survive in the face of cataclysmic astrophysical events, such as asteroid impacts, supernovae, or gamma-ray bursts. This rather extraordinary conclusion was based primarily on the remarkable physiological adaptations and tolerances to extreme conditions displayed by tardigrades1 — those gloriously cute, but tiny (most are around 0.5 mm long as adults) ‘water bears’ or ‘moss piglets’ — could you get any cuter names?
Found almost everywhere and always (the first fossils of them date back to the early Cambrian over half a billion years ago), these wonderful little creatures are some of the toughest metazoans (multicellular animals) on the planet. Only a few types of extremophile bacteria are tougher.
So, boil, fry or freeze the Earth, and you’ll still have tardigrades around, concluded Sloan and colleagues.
When Giovanni first read this, and then passed the paper along to me for comment, our knee-jerk reaction as ecologists was a resounding ‘bullshit!’. Even neophyte ecologists know intuitively that because species are all interconnected in vast networks linked by trophic (who eats whom), competitive, and other ecological functions (known collectively as ‘multiplex networks’), they cannot be singled out using mere thermal tolerances to predict the probability of annihilation. Read the rest of this entry »
I’m not sure if many South Australians are aware of this, but the Parliamentary Inquiry into Biodiversity by the Environment, Resources and Development Committee presented a report to the 53rd Parliament of South Australia in March 2017. I thought it worthwhile reproducing their executive summary here on CB.com (I’ve highlighted the text that I deem to be rather insightful and simultaneously damning from our own elected government representatives):
—
This report summarises the findings and recommendations of the South Australian Parliament’s Environment, Resources and Development Committee’s inquiry into biodiversity in South Australia. Specifically, the inquiry investigated the regulatory and policy framework to determine whether it appropriately supports terrestrial and marine ecological processes, biodiversity values and abates species extinction.
The Committee found that in spite of the efforts of the State and Federal governments, industry and private landholders in South Australia, the condition of biodiversity in the State continues to decline. Species extinctions have occurred in the past and a further “extinction debt” still exists. There is no reason to believe that this trend will improve without a change to the way we approach biodiversity conservation.
A key theme to emerge from the Inquiry is that biodiversity conservation needs to be everyone’s responsibility; State and Federal government, industry, the broader community, and private landholders.
This also means that biodiversity conservation needs to occur across both public and private land, with actions coordinated at a landscape scale.
Making biodiversity conservation everyone’s responsibility requires a range of measures, including legislative reform, improved management of threats and greater involvement of the community. The provision of greater resources would yield faster results.
This report has focused on several key themes that emerged from submissions to the Inquiry.
Regulating for better biodiversity – South Australia’s legislative framework
South Australia’s current legislative framework does not provide for optimum biodiversity outcomes.
Three key issues contribute to this –
an out-of-date suite of environmental legislation that lacks cohesion and consistency, particularly regarding enforcement and compliance provisions;
inadequate and incomplete processes for identifying and protecting at-risk elements that need special measures (e.g. for protection of specific threatened species and ecological communities); and
inadequate consideration of biodiversity conservation in legislation that regulates human activities. In particular, there is a lack of cohesion between the environmental legislative and policy framework and land use planning, assessment and approval.
Statutory fragmentation of biodiversity considerations – that is, consideration of different aspects of biodiversity under different pieces of legislation – results in lack of cohesion and consistency, duplication and inefficiency, and makes it difficult to implement a landscape approach or to identify strategic opportunities and risks.
Taken as a whole, current enforcement provisions do not provide for effective and proportionate compliance action. Enforcement and compliance provisions across the relevant legislation are uneven in their approach. For example, penalties appear to be disproportionate and not risk-based (although there are some exceptions). Modern enforcement tools such as compliance orders, civil remedies and alternative penalties (such as administrative penalties, payment of damages including exemplary damages, remediation orders etc) are not included in all relevant legislation. There is some duplication in offences and inconsistency in the types of sanctions and penalty ranges.
There is an urgent need to amend the legislative framework to support any attempt to improve biodiversity outcomes.
The best approach will be based on clear, shared responsibility for biodiversity outcomes, supported by individual accountability. However, such a change will require policy development and drive.
To ensure forward momentum and improvements in the short term while developing the policy settings to support such a step-change, a staged approach could be implemented. There are various ways this could be achieved.
The Committee suggests a 3-stage approach to reforming the legislative framework. The Committee recommends the creation of a Biodiversity Expert Panel that is responsible for advancing this 3-stage approach.
The first stage will involve amendments to improve operation and effectiveness of the regulatory regime within current policy settings, acknowledging that as a result of Stage 3, provisions may be altered or moved into different pieces of legislation. Amendments generally would be to the existing ‘environmental’ Acts, and primarily to the National Parks and Wildlife Act 1972 and Native Vegetation Act 1991. They would include many of the specific areas for amendment identified in EDO submissions (2011 & 2015) as well as in the SA Government submission, for example, beginning with amendments to improve current environmental legislation.
Stage 2 would progress to amendments to improve integration between Acts and improve support for landholders and community participation.
Stage 3 would implement a system whereby all resource use and management would be managed by one piece of legislation, with protection of biodiversity and sustainable development at its core. Provisions for protected area management, and for the scientific work involved in identifying threatened species and communities, may be contained in separate legislation.
Threats, ecological resilience and restoration
The State’s native biodiversity is facing myriad of current threats, including habitat loss and fragmentation (due to development and changing land-use), pest plants and animals, and control burn regimes. There is a need for more stringent vegetation protection, better informed and enacted control and management strategies of known pest plants and animals, and a revision of burning regimes.
Future threats to the State’s biodiversity will be largely driven by climate change impacts and the interaction with existing major threats (e.g. urbanisation and changing land use). Adequately preparing for and managing such future threats will require knowledge of projected changes and pro-active preparation for such changes.
Working with the community
Involvement of the community is an essential part of any biodiversity conservation strategy for the State. It is a foundation stone for moving to a point where biodiversity conservation is everyone’s business.
Community engagement will become increasingly important for biodiversity conservation, especially given the growing role of volunteers to support works on public land as well as the voluntary conservation efforts of private landholders. The expanding role of volunteers reenforces that biodiversity conservation is everyone’s business.
South Australia’s approach to biodiversity conversation on private land needs to be reinvigorated.
Cross cutting themes
There were several cross cutting themes identified in submissions to the Inquiry. There was broad recognition of the strong cultural and historic significance of elements of biodiversity to Aboriginal people, and that this is often poorly understood outside those communities. Continuing to identify ways for Aboriginal people to contribute to land and water management in South Australia remains a priority.
With respect to knowledge generation, critical knowledge gaps exist that need to be filled and existing knowledge is not being adequately understood, communicated or applied. From a resourcing perspective, there is concern that insufficient funds are being allocated to biodiversity conservation, which is affecting work on public and private lands.
The management of over-abundant species in South Australia remains a challenge, noting the recent impacts of long-nose fur seals in the Lower Lakes and Coorong, and ongoing concerns regarding the impact of animals such as little corellas and some species of kangaroos on negative vegetation.
I published this last week on The Conversation, and now reproducing it here for CB.com readers.
—
Two days ago, the last male northern white rhino (Ceratotherium simum cottoni) died. His passing leaves two surviving members of his subspecies: both females who are unable to bear calves.
Even though it might not be quite the end of the northern white rhino because of the possibility of implanting frozen embryos in their southern cousins (C. simum simum), in practical terms, it nevertheless represents the end of a long decline for the subspecies. It also raises the question: how many individuals does a species need to persist?
Fiction writers have enthusiastically embraced this question, most often in the post-apocalypsegenre. It’s a notion with a long past; the Adam and Eve myth is of course based on a single breeding pair populating the entire world, as is the case described in the Ragnarok, the final battle of the gods in Norse mythology.
This idea dovetails neatly with the image of Noah’s animals marching “two by two” into the Ark. But the science of “minimum viable populations” tells us a different story.
I’ve just read an excellent paper that succinctly, eloquently, and wisely summarised the current predicament of our highly interconnected, global, complex adaptive system (i.e., our environment).
If you are new to the discussions around state shifts, hysteresis, tipping points, and system collapse, there might be a lot in the new paper by Philip Garnett of the University of York that you could find intimidating (and not just because of the complexity of the concepts he discusses). If you are more up-to-date on these discussions, I highly recommend reading this paper for distilling some of the more pertinent questions.
The essence of the paper is that our global environment (Earth) is one giant, complex system made up of interacting sub-systems. We can think of these as a giant, interconnected network of nodes and connections (often called ‘edges’) between them. If you do ecological network theory, then you know what I’m talking about.
What’s particularly fascinating to me is that Philip Garnett is not an environmental scientist; in fact, he’s a a lecturer in Operations Management and Business Analytics (although he does have a background in genetics and biology) who specialises in complex systems theory. In fact, much of his paper uses socio-economic examples of system complexity and collapse, yet the applications to environmentalism in general, and to ecological integrity in particular, are spot on.
As I have done for the last four years (2016, 2015, 2014, 2013), here’s another retrospective list of the top 20 influential conservation papers of 2017 as assessed by experts in F1000 Prime.
I’ve recently read perhaps the most comprehensive treatise of forest fragmentation research ever compiled, and I personally view this rather readable and succinct review by Bill Laurance and colleagues as something every ecology and conservation student should read.
The ‘Biological Dynamics of Forest Fragments Project‘ (BDFFP) is unquestionably one of the most important landscape-scale experiments ever conceived and implemented, now having run 38 years since its inception in 1979. Indeed, it was way ahead of its time.
Experimental studies in ecology are comparatively rare, namely because it is difficult, expensive, and challenging in the extreme to manipulate entire ecosystems to test specific hypotheses relating to the response of biodiversity to environmental change. Thus, we ecologists tend to rely more on mensurative designs that use existing variation in the landscape (or over time) to infer mechanisms of community change. Of course, such experiments have to be large to be meaningful, which is one reason why the 1000 km2 BDFFP has been so successful as the gold standard for determining the effects of forest fragmentation on biodiversity.
With the official start of the new ARCCentre of Excellence for Australian Biodiversity and Heritage (CABAH) in July, I am pleased to announce two new CABAH-funded postdoctoral positions (a.k.a. Research Associates) in my global ecology lab at Flinders University in Adelaide (Flinders Modelling Node).
One of these positions is a little different, and represents something of an experiment. The Research Associate in Palaeo-Vegetation Modelling is being restricted to women candidates; in other words, we’re only accepting applications from women for this one. In a quest to improve the gender balance in my lab and in universities in general, this is a step in the right direction.
The project itself is not overly prescribed, but we would like something along the following lines of inquiry: Read the rest of this entry »
One of the greatest unmet challenges in conservation biology is the genetic management of fragmented populations of threatened animal and plant species. More than a million small, isolated, population fragments of threatened species are likely suffering inbreeding depression, loss of evolutionary potential, and elevated extinction risks (genetic erosion). Re-establishing gene flow between populations is required to reverse these effects, but managers very rarely do this. On the contrary, molecular genetic methods are mainly being used to document genetic differentiation among populations, with most studies concluding that genetically differentiated populations should be managed separately (i.e., kept isolated), thereby dooming many populations to eventual extinction.
The need for a paradigm shift in genetic management of fragmented populations has been highlighted as a major issue in conservation. The rapidly advancing field of molecular genetics is continually providing new tools to measure the extent of population fragmentation and its genetic consequences. However, adequate guidance on how to use these data for effective conservation is still lacking, and many populations are going extinct principally for genetic reasons. Consequently, there is now urgent need for an authoritative textbook on the subject.
Just over two years ago I reported the ‘likely’ eradication of feral pigs from Australia’s third-largest (4,405 km2) island — Kangaroo Island. I indicated ‘likely’ because the program still required the proof-of-eradication phase to be completed before an official declaration could be made. Yesterday I had the immense honour to take part in the official…
Have you ever done any research that relied to any degree on Indigenous Knowledges? How did you cite those Knowledges, if at all? It’s probably time we rethink how we engage with Indigenous Knowledge systems. In a new article published in BioScience, we — a large group of Indigenous and non-Indigenous scholars in Australia —…
A recent paper, co-authored with the late Paul Ehrlich, reveals that the global human population has surpassed Earth’s sustainable capacity. It highlights the dire implications for food security, climate stability, and wellbeing. The study underscores that immediate changes in consumption and population management are crucial for a sustainable future.








 Around a fortnight ago I wrote a hastily penned post about the
Around a fortnight ago I wrote a hastily penned post about the 

 Just under two weeks ago,
Just under two weeks ago, 
 I’m not sure if many South Australians are aware of this, but the Parliamentary Inquiry into Biodiversity by the
I’m not sure if many South Australians are aware of this, but the Parliamentary Inquiry into Biodiversity by the 







