A few weeks ago we published a paper that was in the works for a long time, so long in fact that one of my co-authors died before it was published online.
Paul Ehrlich died on 13 March 2026 at the ripe old age of 93, exactly two weeks before our article appeared online. Paul had a good innings no doubt, but I wish he had survived long enough to see what might very well be his last co-authored paper.
I first met Paul back in the mid-2000s during a trip through San Francisco. I had organised to come chat with Professor Gretchen Daily at Stanford, and Paul came along for lunch. I remember vividly how we clicked almost immediately.
We clicked so well in fact, that we wrote a book together, co-authored several high-impact papers (e.g., ‘ghastly future‘), spent a month in Bellagio as Rockefeller Foundation writing residents, participated in various public and parliamentary presentations, and generally just got on like a house on fire. Paul and his wife Ann became like family, so much so that they were de facto grandparents to my daughter who grew up with them in near-annual contact.
This post isn’t about Paul per se, but I cannot ignore the profound influence Paul had on my career, my personality, and my life view. I miss him. I am therefore dedicating this paper and post to his memory. So long, and thanks for all the fish.
Nothing like writing about human population to get the punters engaged.
We show empirically that the Earth has already exceeded its ability to support the global human population sustainably, with dire implications for increasing pressure on food security, climate stability, and human wellbeing. However, slowing population growth and raising global awareness could still offer us some hope.
Our study shows that humans have pushed well beyond the planet’s long-term carrying capacity and that continued growth under current patterns of consumption will intensify environmental and social challenges for communities worldwide.
In boreal forests, many hares adopt white winter coats before the snow arrives. In a snowless landscape, these white hares lack camouflage against predators. However, their early moult from brown into white fur can increase their survival and offers an advantage as the snow season becomes progressively shorter with climate change.
Throughout the year, we wear different clothing to protect ourselves from the cold or heat and for aesthetic reasons depending on the occasion. Likewise, many animals change the colour, thickness and structure of their fur and feathers in tune with the seasons.
Snowshoe hare (Lepus americanus) in a snowy (Kluane Lake/Yukon, Canada) and snowless habitat (Seely Lake/Montana, USA). This mammal moults its coat as colder temperatures, shorter days, and snowfall arrive. In the genetic populations of the temperate forests of the Rocky Mountains and the boreal forests spanning the North American continent, hares that moult from brown to white are abundant (20). However, in coastal areas, and in the third genetic population in the North Pacific, snowfall is brief and less intense, resulting in fewer white individuals. This is due to hybridisation with the black-tailed jackrabbit (Lepus californicus) over 3,000 years ago (17). The hare’s coat has an outer layer, where the longer fur gives each individual its colour, and an inner layer of short fur (19). In winter, the outer layer becomes thicker and denser, while the inner layer maintains a consistent thickness but increases in density. By biomass, the snowshoe hare is the primary herbivore in the North American boreal forest and distinguishes the trophic relationships between continents (21). In Europe, much of the boreal understory remains under snow, providing food for rodents with four-year abundance cycles controlled by small generalist predators (mustelids). In North America, the boreal understory grows above the snow and provides food for hares. In this region, snowshoe hare populations follow 10-year abundance cycles regulated by specialist predators (those that feed almost exclusively on hares), primarily the Canada lynx (Lynx canadensis) (6). Photos courtesy of Alice Kenney and Charles Krebs (Yukon) [see their ecological monitoring program here] and Marketa Zimova (Montana).
However, as the climate changes, springs arrive earlier, winters are delayed, and the frequency and intensity of precipitation have become highly variable. All of this makes it harder for species to adjust their wardrobe to temperature changes (1).
In this context, body colour is a critical factor for birds and mammals that undergo an annual moult (2). In 21 species from the cold latitudes of the Northern Hemisphere, some individuals are brown in summer, but turn white in winter, while others remain brown year round (3). This phenomenon includes weasels, rodents, ptarmigans, foxes, rabbits and hares.
Coral reefs are much more than just a pretty place to visit. They are among the world’s richest ecosystems, hosting about a third of all marine species.
These reefs also directly benefit more than a billion people, providing livelihoods and food security, as well as protection from storms and coastal erosion.
Without coral reefs, the world would be a much poorer place. So when corals die or become damaged, many people try to restore them. But the enormity of the task is growing as the climate keeps warming.
In our new research, we examined the full extent of existing coral restoration projects worldwide. We looked at what drives their success or failure, and how much it would actually cost to restore what’s already been lost. Restoring the reefs we’ve already lost around the world could cost up to A$26 trillion.
Bleached Acropora corals in the Maldives.Davide Seveso/University of Milan
When sea temperatures climb above the seasonal average for sustained periods, corals can become bleached. They lose colour as they expel their symbiotic algae when stressed, revealing the white skeleton underneath. Severe bleaching can kill coral.
Yes, it’s bad, especially for US-based scientists. It also affects scientists in Australia and the rest of the world. But there are ways to get around the problem. There might even be a silver lining to this dark cloud.
Trump cannot stop global climate action, although he might slow it. Nor can he hide the truth by restricting access to data. Climate research will continue despite Trump’s best efforts to hamstring scientists and research institutions.
No strength in ignorance
Last year was the warmest on record, a fact that yet again confirms our worst-case predictions. The world has already surpassed the (arbitrary) 1.5°C threshold increase relative to pre-industrial temperatures — a threshold that only a few years ago we didn’t think we would cross until 2030 at the earliest.
We’re now on track to be living in a world that’s 3°C hotter or more by the end of the century.
But ignoring climate change won’t make it go away. Like the Ministry of Truth in George Orwell’s classic dystopian novel, 1984, Trump seems to believe “ignorance is strength”. He’s trying to erase facts about the climate crisis, perhaps to keep people ignorant and subdued.
What this means for Australian climate science
Many Australian scientists (including me) collaborate regularly with US colleagues, share funding, and publish results together. Knowledge sharing and open-access data are the foundation of advances in science, so Trump’s assault will inevitably slow progress here.
For example, Australian and US scientists regularly collaborate in big-ticket research and policy development related to climate change, such as the Intergovernmental Panel on Climate Change’s Physical Science Basis reports. But even with fewer US scientists in the mix, the research and reporting will continue.
Other reputable climate-data repositories around the world include the European Union’s Climate Data Store, the University of East Anglia’s Climate Research Unit, the Netherlands Meteorological Institute’s Climate Explorer, and the independent WorldClim, to name a few.
While restricting access to US-based websites is inconvenient, we can readily get around the problem. Many of my colleagues have also been downloading data prior to the purge mandate to maintain access.
Consequences for the US
Over the past month I have been inundated with horror stories from many US-based colleagues in academia and the public service, who have lost their jobs and/or research funding. In addition to these very real personal tragedies, the bigger picture is even bleaker.
The loss of scientific and technical expertise these mass sackings entail weakens the capability of the US workforce to discover and develop solutions to climate change. Just when we need good scientific and engineering innovations more than ever, a massive capacity is being erased before our eyes.
More emissions mean more climate change, especially when you’re already one of the biggest contributors to the global problem. The US is the second-highest greenhouse emitter in the world, behind only China.
On his first day as president, Trump withdrew the US from the Paris climate agreement. This effectively removes his country from all binding limits on actions that contribute to climate change.
Weakening international treaties is a two-edged sword, because it not only lets the US off the leash, it also potentially discourages other nations from acting responsibly. Analogous to the “unresponsive bystander effect”, many nations may now be more hesitant to commit to reductions because one of the biggest emitters refuses to do anything about it.
Trump has also slashed US international aid, which will slow climate action in countries that need the most assistance.
Overall, faster rates of warming will inevitably put more strain on natural resources and agricultural production. This could increase the probability of international warfare over water, food and other essential natural resources. Because autocratic countries cope worse with food shortages than democratic ones, climate emergencies will penalise nations led by despots more heavily.
Trump’s foolhardy anti-climate campaign is enough to make many people despair. But there are a few faint glimmers of hope on the horizon.
As the US shirks its domestic and international responsibilities, other countries might resolve to do more. Not relying on the US could force capacity-building elsewhere. Some even suggest without the US at the table slowing progress, stronger climate action might result.
Americans have their own daunting fight on their hands. But the rest of the world will have to take up the slack if we have any chance of limiting the health, wealth, equality, human rights and biodiversity calamities now unfolding because of climate change.
Corey J. A. Bradshaw, Matthew Flinders Professor of Global Ecology and Node Leader in the ARC Centre of Excellence for Indigenous and Environmental Histories and Futures, Flinders University
Human overpopulation is often depicted in the media in one of two ways: as either a catastrophic disaster or an overly-exaggerated concern. Yet the data understood by scientists and researchers is clear. So what is the actual state of our overshoot, and, despite our growing numbers, are we already seeing the signs that the sixth mass extinction is underway?
In a recent episode of The Great Simplification podcast, Nate Hagens was joined by global ecologist Corey Bradshaw to discuss his recent research on the rapid decline in biodiversity, how population and demographics will change in the coming decades, and what both of these will mean for complex global economies currently reliant on a stable environment.
In Australia, most fire occurs in the vast tropical savannas of the country’s north. In new research published in Nature Geoscience, we show Indigenous management of fire in these regions began at least 11,000 years ago – and possibly as long as 40,000 years ago.
But climate change and other effects of human activity are making wildfires more common and more severe in many regions, often with catastrophic results. In Australia, fires have caused major economic, environmental and personal losses, most recently in the south of the country.
Australia is home to about one in 12 of the world’s species of animals, birds, plants and insects – between 600,000 and 700,000 species. More than 80% of Australian plants and mammals and just under 50% of our birds are found nowhere else.
But habitat destruction, climate change, and invasive species are wreaking havoc on Earth’s rich biodiversity, and Australia is no exception.
More and more species stand on the edge of oblivion. That’s just the ones we know enough about to list formally as threatened. Many more are in trouble, especially in the oceans. Change is the new constant. As the world heats up and ecosystems warp, new combinations of species can emerge without an evolutionary connection, creating novel communities.
It is still possible to stop species from dying out. But it will take an unprecedented effort.
The vulnerable southern bell (growling grass) frog (Litoria raniformis). Rupert Mathwin/Flinders University
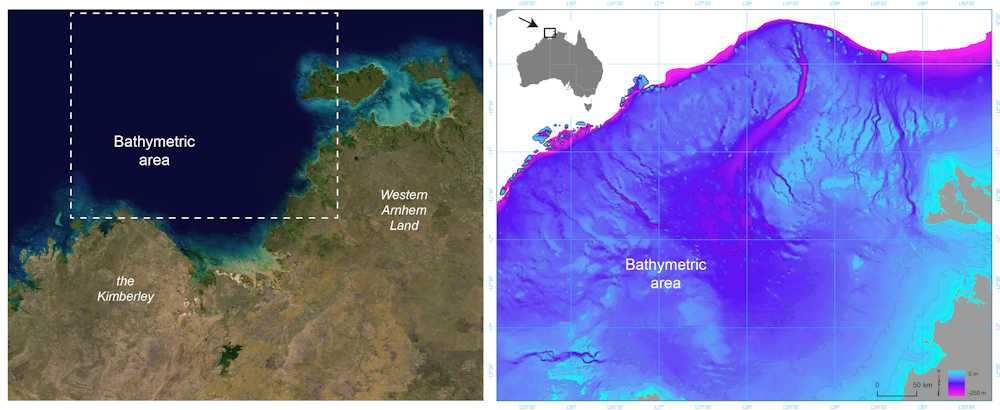
For much of the 65,000 years of Australia’s human history, the now-submerged northwest continental shelf connected the Kimberley and western Arnhem Land. This vast, habitable realm covered nearly 390,000 square kilometres, an area one-and-a-half times larger than New Zealand is today.
Left: Satellite image of the submerged northwest shelf region. Right: Drowned landscape map of the study area. US Geological Survey, Geoscience Australia
It was likely a single cultural zone, with similarities in ground stone-axe technology, styles of rock art, and languages found by archaeologists in the Kimberley and Arnhem Land.
There is plenty of archaeological evidence humans once lived on continental shelves – areas that are now submerged – all around the world. Such hard evidence has been retrieved from underwater sites in the North Sea, Baltic Sea and Mediterranean Sea, and along the coasts of North and South America, South Africa and Australia.
In a newly published study in Quaternary Science Reviews, we reveal details of the complex landscape that existed on the Northwest Shelf of Australia. It was unlike any landscape found on our continent today.
A continental split
Around 18,000 years ago, the last ice age ended. Subsequent warming caused sea levels to rise and drown huge areas of the world’s continents. This process split the supercontinent of Sahul into New Guinea and Australia, and cut Tasmania off from the mainland.
Unlike in the rest of the world, the now-drowned continental shelves of Australia were thought to be environmentally unproductive and little used by First Nations peoples.
But mounting archaeological evidence shows this assumption is incorrect. Many large islands off Australia’s coast – islands that once formed part of the continental shelves – show signs of occupation before sea levels rose.
Stone tools have also recently been found on the sea floor off the coast of the Pilbara region of Western Australia.
A global database set up by scientists to assemble data on the economic cost of biological invasions in support of effective government management strategies has grown to include all known invasive species.
Now involving 145 researchers from 44 countries — the current version of InvaCost has 13,553 entries in 22 languages and enables scientists to develop a clear picture about the major threats globally of invasive species to ecosystems, biodiversity, and human well-being.
Biological invasions are caused by species introduced on purpose or accidentally by humans to areas outside of their natural ranges. From cats and weeds, to crop pests and diseases, invasive species are a worldwide scourge.
Invasive species have cost over US$2 trillion globally since the 1970s by damaging goods and services, and through the costs of managing them, and these economic costs are only increasing.
A new synthesis published in the journal BioSciencedocuments the progress of the InvaCost endeavour.The study provides a timeline of the state of invasion costs, starting with prior flaws and shortcomings in the scientific literature, then how InvaCost has helped to alleviate and address these issues, and what the future potentially holds for research and policymakers.
But just how far-fetched is it? Could pathogens that were once common on Earth – but frozen for millennia in glaciers, ice caps and permafrost – emerge from the melting ice to lay waste to modern ecosystems? The potential is, in fact, quite real.
In 2014, a giant “zombie” Pithovirus sibericum virus was revived from 30,000-year-old Siberian permafrost.
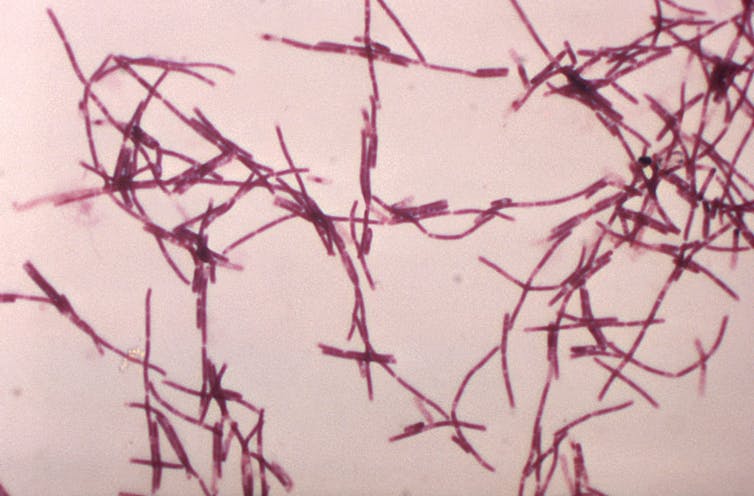
And in 2016, an outbreak of anthrax (a disease caused by the bacterium Bacillus anthracis) in western Siberia was attributed to the rapid thawing of B. anthracis spores in permafrost. It killed thousands of reindeer and affected dozens of people.
Bacillus anthracis is a soil bacterium that causes anthrax. William A. Clark/USCDCP
More recently, scientists found remarkable genetic compatibility between viruses isolated from lake sediments in the high Arctic and potential living hosts.
Earth’s climate is warming at a spectacular rate, and up to four times faster in colder regions such as the Arctic. Estimates suggest we can expect four sextillion (4,000,000,000,000,000,000,000) microorganisms to be released from ice melt each year. This is about the same as the estimated number of stars in the universe.
However, despite the unfathomably large number of microorganisms being released from melting ice (including pathogens that can potentially infect modern species), no one has been able to estimate the risk this poses to modern ecosystems.
In a new study published today in the journal PLOS Computational Biology, we calculated the ecological risks posed by the release of unpredictable ancient viruses.
Have you ever watched a nature documentary and marvelled at the intricate dance of life unfolding on screen? From the smallest insect to the largest predator, every creature plays a role in the grand performance of our planet’s biosphere. But what happens when one of these performers disappears?
In this post, we delve into our recent article Estimating co-extinction risks in terrestrial ecosystems just published in Global Change Biology, in which we discuss the cascading effects of species loss and the risks of ‘co-extinction’.
But what does ‘co-extinction’ really mean?
Imagine an ecosystem as a giant web of interconnected species. Each thread represents a relationship between two species — for example, a bird that eats a certain type of insect, or a plant that relies on a specific species of bee for pollination. Now, what happens if one of these species in the pair disappears? The thread breaks and the remaining species loses an interaction. This could potentially lead to its co-extinction, which is essentially the domino effect of multiple species losses in an ecosystem.
A famous example of this effect can be seen with the invasion of the cane toad (Rhinella marina) across mainland Australia, which have caused trophic cascades and species compositional changes in these communities.
The direct extinction of one species, caused by effects such as global warming for example, has the potential to cause other species also to become extinct indirectly.
Wildfires transform forests into mosaics of vegetation. What, where, and which plants thrive depends on when and how severely a fire affects different areas of a forest. Such heterogeneity in the landscape is essential for animal species that benefit from fire like woodpeckers.
The black-backed woodpecker (Picoides arcticus) lives in the coniferous forests of North America’s boreal-Mediterranean region. Thanks to a powerful and sharp bill, this bird can excavate nests inside the trunks of (mainly dead) trees, and those cavities will be re-used later by many species of birds, mammals, and invertebrates in fire-prone landscapes (22). The images show a male with the characteristic black plumage of his back that serves as camouflage against the dark bark of a dead tree three years after a wildfire in Montana (USA). Being omnivores, the diet of this bird largely relies on the larvae of woodboring coleoptera like jewell and longhorn beetles. These insects are abundant post-fire, the champion being the fire beetle (Melanophila spp.). The thorax of fire beetles is equipped with infrared-light receptors that can detect a wildfire from tens of kilometres away (23). These fascinating little beasts are the first to arrive at a burned forest and, of course, woodpeckers follow soon after. The preference of the blackbacked woodpecker for burned forests and their cryptic feathers and pyrophilic diet reflect a long evolutionary history in response to fires. Courtesy of Richard Hutto.
Anyone raised in rural areas will have vivid recollections of wildfires: the thick, ashy smell, the overcast sky on a sunny day, and the purring of aerial firefighters dropping water from their hanging tanks. The reality is that wildfires are natural events that shape biodiversity and ecosystem function (1) — to the extent that fire is intimately linked to the appearance and evolution of terrestrial plants (2). Since the Palaeolithic, our own species has used fire at will, to cook, hunt, melt metals, open cropland or paths, or tell stories in front of a hearth (3).
Where there are regular wildfires (fire-prone ecosystems), different areas of the landscape burn in different seasons and years under different weather patterns. Therefore, each region has a unique fire biography in terms of how frequently, how much, and how long ago wildfires occurred. All those factors interact will one another and with topography.
Each organism has a limit of tolerance to cold and hot temperatures. So, the closer it lives to those limits, the higher the chances of experiencing thermal stress and potentially dying. In our recent paper, we revise gaps in the knowledge of tolerance to high temperatures in cold-blooded animals (ectotherms), a diverse group mostly including amphibians and reptiles (> 16,000 species), fish (> 34,000 species), and invertebrates (> 1,200,000 species).
As a scientist, little is more self-realising than to write and publish a conceptual paper that frames the findings of your own previous applied-research papers. This is the case with an opinion piece we have just published in Basic and Applied Ecology1 — 10 years, 4 research papers2-5 [see related blog posts here, here, here and here], and 1 popular-science article6 after I joined the Department of Biogeography and Global Change (Spanish National Research Council) to study the thermal physiology of Iberian lizards under the supervision of Miguel Araújo and David Vieites.
Iberian lizards for which heat tolerance is known (varying from 40 to 45 °C)
[left, top to bottom] Iberian emerald lizard (Lacerta schreiberi, from Alameda del Valle/Madrid) and Geniez’s wall lizard (Podarcis virescens, Fuertescusa/Cuenca), and [right, top to bottom] Algerian sand racer (Psammodromus algirus, Navacerrada/Madrid), Andalusian wall lizard (Podarcis vaucheri, La Barrosa/Cádiz), Valverde’s lizard (Algyroides marchi, Riópar/Albacete), and Cyren’s rock lizard (Iberolacerta cyreni, Valdesquí/Madrid). Heat-tolerance data deposited here and used to evaluate instraspecific variation of heat tolerance3,4. Photos: Salvador Herrando-Pérez.
In our new paper, we examine how much we know and what areas of research require further development to advance our understanding of how and why the tolerance of ectotherm fauna to high environmental temperature (‘heat tolerance’ hereafter) varies within and across the Earth’s biomes. We focus on data gaps using the global database GlobTherm as a reference template (see Box 1 below).
Our three main tenets
1. Population versus species data: Most large-scale ecophysiological research is based on modelling one measurement of heat tolerance per species (typically representing one population and/or physiological assay) over hundreds to thousands of species covering broad geographical, phylogenetic, and climatic gradients.
But there is ample evidence that heat tolerance changes a lot among populations occupying different areas of the distribution of a species, and such variation must be taken into account to improve our predictions of how species might respond to environmental change and face extinction.
Flooding in the Murray-Darling Basin is creating ideal breeding conditions for many native species that have evolved to take advantage of temporary flood conditions. Led by PhD candidate Rupert Mathwin, our team developed virtual models of the Murray River to reveal a crucial link between natural flooding and the extinction risk of endangered southern bell frogs (Litoria raniformis; also known as growling grass frogs).
Southern bell frogs are one of Australia’s 100 Priority Threatened Species. This endangered frog breeds during spring and summer when water levels increase in their wetlands. However, the natural flooding patterns in Australia’s largest river system have been negatively impacted by expansive river regulation that some years, sees up to 60% of river water extracted for human use.
Our latest paper describes how we built computer simulations of Murray-Darling Basin wetlands filled with simulated southern bell frogs. By changing the simulation from natural to regulated conditions, we showed that modern conditions dramatically increase the extinction risk of these beloved frogs.
The data clearly indicate that successive dry years raise the probability of local extinction, and these effects are strongest in smaller wetlands. Larger wetlands and those with more frequent inundation are less prone to these effects, although they are not immune to them entirely. The models present a warning — we have greatly modified the way the river behaves, and the modern river cannot support the long-term survival of southern bell frogs.’
Following my annual tradition, I present the retrospective list of the ‘top’ 20 influential papers of 2022 as assessed by experts in Faculty Opinions(formerly known as F1000). These are in no particular order. See previous years’ lists here: 2021, 2020, 2019, 2018, 2017, 2016, 2015, 2014, and 2013.
Climate change is one of the main drivers of species loss globally. We know more plants and animals will die as heatwaves, bushfires, droughts and other natural disasters worsen.
But to date, science has vastly underestimated the true toll climate change and habitat destruction will have on biodiversity. That’s because it has largely neglected to consider the extent of “co-extinctions”: when species go extinct because other species on which they depend die out.
Our new research shows 10% of land animals could disappear from particular geographic areas by 2050, and almost 30% by 2100. This is more than double previous predictions. It means children born today who live to their 70s will witness literally thousands of animals disappear in their lifetime, from lizards and frogs to iconic mammals such as elephants and koalas.
But if we manage to dramatically reduce carbon emissions globally, we could save thousands of species from local extinction this century alone.
Ravages of drought will only worsen in coming decades. CJA Bradshaw
An extinction crisis unfolding
Every species depends on others in some way. So when a species dies out, the repercussions can ripple through an ecosystem.
For example, consider what happens when a species goes extinct due to a disturbance such as habitat loss. This is known as a “primary” extinction. It can then mean a predator loses its prey, a parasite loses its host or a flowering plant loses its pollinators.
A real-life example of a co-extinction that could occur soon is the potential loss of the critically endangered mountain pygmy possum (Burramys parvus) in Australia. Drought, habitat loss, and other pressures have caused the rapid decline of its primary prey, the bogong moth (Agrotis infusa).
The conservation, environment, and sustainability literature is rife with the term ‘collapse’, applied to concepts as diverse as species extinction to the complete breakdown of civilisation. I have also struggled with its various meanings and implications, so I’m going to attempt to provide some clarity on collapse for my own and hopefully some others’ benefit.
From a strictly ecological perspective, ‘collapse’ could be described in the following (paraphrased) ways:
abrupt transition of one ecosystem state to another, usually invoking the idea that something has declined in the process (species richness, beta diversity, functional diversity, trophic network connectance, trait volume, production, etc.);
But there is still nor formal definition of ‘collapse’ in ecology, as identified by several researchers (Keith et al. 2013; Boitani et al. 2015; Keith et al. 2015; Sato and Lindenmayer 2017; Bland et al. 2018). While this oversight has been discussed extensively with respect to quantifying changes, I can find nothing in the literature that attempts a generalisable definition of what collapse should mean. Perhaps this is because it is not possible to identify a definition that is sufficiently generalisable, something that Boitani et al. (2015) described with this statement:
“The definition of collapse is so vague that in practice it will be possible (and often necessary) to define collapse separately for each ecosystem, using a variety of attributes and threshold values
Despite all the work that has occurred since then, I fear we haven’t moved much beyond that conclusion.
Hell, cutting down the trees in the bush block next to my property constitutes a wholesale ‘collapse’ of the microcommunity of species using that patch of bush. An asteroid hitting the Earth and causing a mass extinction is also collapse. And everything in-between.
But at least ecologists have made some attempts to define and quantify collapse, even if an acceptable definition has not been forthcoming. The sustainability and broader environment literature has not even done that.
In a newly announced partnership with Texas biotech company Colossal Biosciences, Australian researchers are hoping their dream to bring back the extinct thylacine is a “giant leap” closer to fruition.
Scientists at University of Melbourne’s TIGRR Lab (Thylacine Integrated Genetic Restoration Research) believe the new partnership, which brings Colossal’s expertise in CRISPR gene editing on board, could result in the first baby thylacine within a decade.
The genetic engineering firm made headlines in 2021 with the announcement of an ambitious plan to bring back something akin to the woolly mammoth, by producing elephant-mammoth hybrids or “mammophants”.
But de-extinction, as this type of research is known, is a highly controversial field. It’s often criticised for attempts at “playing God” or drawing attention away from the conservation of living species. So, should we bring back the thylacine? We asked five experts.
As is my tendency, I like to wade carefully into other disciplines from time to time to examine what components they can bring to the conservation table. I do not profess any sort of expertise when I do so, but if I require a true expert for research purposes, then I will collaborate with said experts.
I often say to my students that in many ways, the science of sustainability and conservation is more or less resolved — what we need now is ways to manage the human side of the problems we face. The disciplines that deal with human management, such as psychology, economics, political science, and sociology, are mainly pursuits of the humanities (have I just argued myself out of a job?).
On the topic of human psychology, I think most people involved in some way with biodiversity conservation often contemplate why human societies are so self-destructive. Even in the face of logic and evidence, people deny what’s going on in front of their eyes (think anti-vaxxers, climate-change denialists, etc.), so it should be no wonder why many (most?) people deny their own existential threats. Yet, it still doesn’t seem to make much sense to us until we put the phenomenon into a psychological framework.
My apologies here to actual psychologists if I oversimplify or otherwise make mistakes, but the following explanation has done a lot for me personally in my own journey to understand this conundrum. It is also a good way to teach others about why there is so much reticence to fixing our environmental problems.
The idea is a rather simple one, but it requires a little journey to appreciate. Let’s pop back to the 1970s with the publication of Ernest Becker’s The Denial of Death, for which he won the Pulitzer Prize in 1974 (ironically, two months after his own death). In this book, Becker examined the awareness of death on human behaviour and the strategies that we have developed to mitigate our fear of it. This particular quote sums it up nicely:
This is the terror: to have emerged from nothing, to have a name, consciousness of self, deep inner feelings, and excruciating inner yearning for life and self expression — and with all this yet to die
Ernest Becker in The Denial of Death (1973)
The upshot is that we have evolved a whole raft of coping mechanisms to this personal existential dread. Some engage in overly hedonic pursuits to numb the anxiety; others try to “tranquillise themselves with the trivial”, essentially ignoring the terror, while others still manage the dread through religion and the hope of an existence beyond the mortal.
Get Conservation Bytes by Email Enter your email address to receive new posts on conservation, biodiversity, and environmental science directly in your inbox. No need to visit the website regularly, and you can unsubscribe at any time.
Hulton Archive / Getty Images We warn readers that the content of this study is confronting and may be distressing. In April 1789, the first smallpox epidemic among Aboriginal people in the Sydney region began, just 16 months after Europeans arrived on the First Fleet. Little is known of the true scale of the epidemic,…
Just over two years ago I reported the ‘likely’ eradication of feral pigs from Australia’s third-largest (4,405 km2) island — Kangaroo Island. I indicated ‘likely’ because the program still required the proof-of-eradication phase to be completed before an official declaration could be made. Yesterday I had the immense honour to take part in the official…
Have you ever done any research that relied to any degree on Indigenous Knowledges? How did you cite those Knowledges, if at all? It’s probably time we rethink how we engage with Indigenous Knowledge systems. In a new article published in BioScience, we — a large group of Indigenous and non-Indigenous scholars in Australia —…