In a newly announced partnership with Texas biotech company Colossal Biosciences, Australian researchers are hoping their dream to bring back the extinct thylacine is a “giant leap” closer to fruition.
Scientists at University of Melbourne’s TIGRR Lab (Thylacine Integrated Genetic Restoration Research) believe the new partnership, which brings Colossal’s expertise in CRISPR gene editing on board, could result in the first baby thylacine within a decade.
The genetic engineering firm made headlines in 2021 with the announcement of an ambitious plan to bring back something akin to the woolly mammoth, by producing elephant-mammoth hybrids or “mammophants”.
But de-extinction, as this type of research is known, is a highly controversial field. It’s often criticised for attempts at “playing God” or drawing attention away from the conservation of living species. So, should we bring back the thylacine? We asked five experts.
Last week, researchers at the University of Melbourne announced that thylacines or Tasmanian tigers, the Australian marsupial predators extinct since the 1930s, could one day be ushered back to life.
The thylacine (Thylacinus cynocephalus), also known as the ‘Tasmanian tiger’ (it was neither Tasmanian, because it was once common in mainland Australia, nor was it related to the tiger), went extinct in Tasmania in the 1930s from persecution by farmers and habitat loss. Art by Eleanor (Nellie) Pease, University of Queensland. Centre of Excellence for Australian Biodiversity and Heritage
Advances in mapping the genome of the thylacine and its living relative the numbat have made the prospect of re-animating the species seem real. As an ecologist, I would personally relish the opportunity to see a living specimen.
The announcement led to some overhyped headlines about the imminent resurrection of the species. But the idea of “de-extinction” faces a variety of technical, ethical and ecological challenges. Critics (like myself) argue it diverts attention and resources from the urgent and achievable task of preventing still-living species from becoming extinct.
The rebirth of the bucardo
The idea of de-extinction goes back at least to the the creation of the San Diego Frozen Zoo in the early 1970s. This project aimed to freeze blood, DNA, tissue, cells, eggs and sperm from exotic and endangered species in the hope of one day recreating them.
The notion gained broad public attention with the first of the Jurassic Park films in 1993. The famous cloning of Dolly the sheep reported in 1996 created a sense that the necessary know-how wasn’t too far off.
The next technological leap came in 2008, with the cloning of a dead mouse that had been frozen at –20℃ for 16 years. If frozen individuals could be cloned, re-animation of a whole species seemed possible.
After this achievement, de-extinction began to look like a potential way to tackle the modern global extinction crisis.
For many years I’ve been interested in modelling the extinction dynamics of megafauna. Apart from co-authoring a few demographically simplified (or largely demographically free) models about how megafauna species could have gone extinct, I have never really tried to capture the full nuances of long-extinct species within a fully structured demographic framework.
That is, until now.
But how do you get the life-history data of an extinct animal that was never directly measured. Surely, things like survival, reproductive output, longevity and even environmental carrying capacity are impossible to discern, and aren’t these necessary for a stage-structured demographic model?
The answer to the first part of that question “it’s possible”, and to the second, it’s “yes”. The most important bit of information we palaeo modellers need to construct something that’s ecologically plausible for an extinct species is an estimate of body mass. Thankfully, palaeontologists are very good at estimating the mass of the things they dig up (with the associated caveats, of course). From such estimates, we can reconstruct everything from equilibrium densities, maximum rate of population growth, age at first breeding, and longevity.
But it’s more complicated than that, of course. In Australia anyway, we’re largely dealing with marsupials (and some monotremes), and they have a rather different life-history mode than most placentals. We therefore have to ‘correct’ the life-history estimates derived from living placental species. Thankfully, evolutionary biologists and ecologists have ways to do that too.
The Pleistocene kangaroo Procoptodon goliah, the largest and most heavily built of the short-faced kangaroos, was the largest and most heavily built kangaroo known. It had an unusually short, flat face and forwardly directed eyes, with a single large toe on each foot (reduced from the more normal count of four). Each forelimb had two long, clawed fingers that would have been used to bring leafy branches within reach.
So with a battery of ecological, demographic, and evolutionary tools, we can now create reasonable stochastic-demographic models for long-gone species, like wombat-like creatures as big as cars, birds more than two metres tall, and lizards more than seven metres long that once roamed the Australian continent.
Ancient clues, in the shape of fossils and archaeological evidence of varying quality scattered across Australia, have formed the basis of several hypotheses about the fate of megafauna that vanished during a peak about 42,000 years ago from the ancient continent of Sahul, comprising mainland Australia, Tasmania, New Guinea and neighbouring islands.
There is a growing consensus that multiple factors were at play, including climate change, the impact of people on the environment, and access to freshwater sources.
Just published in the open-access journal eLife, our latest CABAH paper applies these approaches to assess how susceptible different species were to extinction – and what it means for the survival of species today.
Using various characteristics such as body size, weight, lifespan, survival rate, and fertility, we (Chris Johnson, John Llewelyn, Vera Weisbecker, Giovanni Strona, Frédérik Saltré & me) created population simulation models to predict the likelihood of these species surviving under different types of environmental disturbance.
We compared the results to what we know about the timing of extinction for different megafauna species derived from dated fossil records. We expected to confirm that the most extinction-prone species were the first species to go extinct – but that wasn’t necessarily the case.
While we did find that slower-growing species with lower fertility, like the rhino-sized wombat relative Diprotodon, were generally more susceptible to extinction than more-fecund species like the marsupial ‘tiger’ thylacine, the relative susceptibility rank across species did not match the timing of their extinctions recorded in the fossil record.
Indeed, we found no clear relationship between a species’ inherent vulnerability to extinction — such as being slower and heavier and/or slower to reproduce — and the timing of its extinction in the fossil record.
In fact, we found that most of the living species used for comparison — such as short-beaked echidnas, emus, brush turkeys, and common wombats — were more susceptible on average than their now-extinct counterparts.
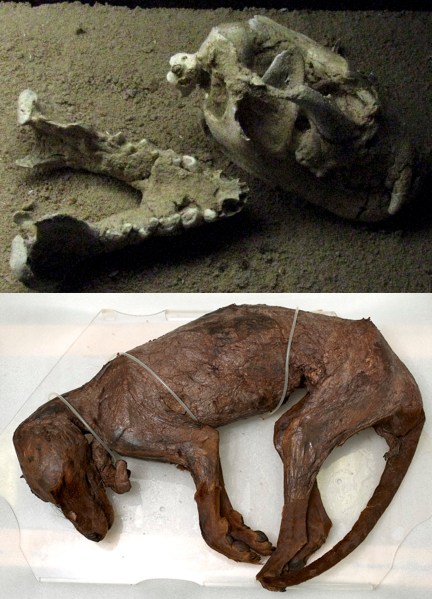
We’ve just published an analysis of new radiocarbon dates showing that thylacines (Tasmanian ‘tigers’, Thylacinus cynocephalus) and Tasmanian devils (Sarcophilus harrisi) went extinct on the Australian mainland at the same time — some 3200 years ago.
For many years, we’ve been uncertain about when thylacines and devils went extinct in mainland Australia (of course, devils are still in Tasmania, and thylacines went extinct there in the 1930s) — a recent age for the devil extinction (500 years before present) has recently been shown to be unreliable. The next youngest reliable devil fossil is 25000 years old.
So, knowing when both species went extinct is essential to be able to determine the drivers of these extinctions, and why they survived in Tasmania. If the two extinctions on the mainland happened at the same time, this would support the hypothesis that a common driver (or set of drivers) caused both species to go extinct. Read the rest of this entry »
Our postdoc, Tom Prowse, has just had one of the slickest set of reviews I’ve ever seen, followed by a quick acceptance of what I think is a pretty sexy paper. Earlier this year his paper in Journal of Animal Ecology showed that thylacine (the badly named ‘Tasmanian tiger‘) was most likely not the victim of some unobserved mystery disease, but instead succumbed to what many large predators have/will: human beings. His latest effort now online in Ecology shows that the thylacine and devil extinctions on the Australian mainland were similarly the result of humans and not the scapegoat dingo. But I’ll let him explain:
—
‘Regime shifts’ can occur in ecosystems when sometimes even a single component is added or changed. Such additions, of say a new predator, or changes such as a rise in temperature, can fundamentally alter core ecosystem functions and processes, causing the ecosystem to switch to some alternative stable state.
Some of the most striking examples of ecological regime shifts are the mass extinctions of large mammals (‘megafauna’) during human prehistory. In Australia, human arrival and subsequent hunting pressure is implicated in the rapid extinction of about 50 mammal species by around 45 thousand years ago. The ensuing alternative stable state was comprised of a reduced diversity of predators, dominated by humans and two native marsupial predators ‑ the thylacine (also known as the marsupial ‘tiger’ or ‘wolf’) and the devil (which is now restricted to Tasmania and threatened by a debilitating, infectious cancer).
Both thylacines and devils lasted on mainland Australia for over 40 thousand years following the arrival of humans. However, a second regime shift resulted in the extinction of both these predators by about 3 thousand years ago, which was coincidentally just after dingoes were introduced to Australia. Dingoes are descended from early domestic dogs and were introduced to northern Australia from Asia by ancient traders approximately 4 thousand years ago. Today, they are Australia’s only top predator remaining, other than invasive European foxes and feral cats. Since the earliest days of European settlement, dingoes have been persecuted because they prey on livestock. During the 1880s, 5614 km of ‘dingo fence’ was constructed to protect south-east Australia’s grazing rangelands from dingo incursions. The fence is maintained to this day, and dingoes are poisoned and shot both inside and outside this barrier, despite mounting evidence that these predators play a key role in maintaining native ecosystems, largely by suppressing invasive predators.
Perhaps because the public perception of dingoes as ‘sheep-killers’ is so firmly entrenched, it has been commonly assumed that dingoes killed off the thylacines and devils on mainland Australia. People who support this view also point out that thylacines and devils persisted on the island of Tasmania, which was never colonised by dingoes (although thylacines went extinct there too in the early 1900s). To date, most discussion of the mainland thylacine and devil extinctions has focused on the possibility that dingoes disrupted the system by ‘exploitation competition’ (eating the same prey), ‘interference competition’ (wasting the native predators’ precious munching time), as well as ‘direct predation’ (dingoes actually eating devils and thylacines). Read the rest of this entry »
If you’re a rational person driven by evidence rather than hearsay, vested interest or faith, then the above examples probably sound ridiculous. But rest assured, millions of people adhere to these points of view because of the phenomenon mentioned in the first sentence above. With this background then, I introduce a paper that’s almost available online (i.e., we have the DOI, but the online version is yet to appear). Produced by our extremely clever post-doc, Tom Prowse, the paper is entitled: No need for disease: testing extinction hypotheses for the thylacine using multispecies metamodels, and will soon appear in Journal of Animal Ecology.
Of course, I am biased being a co-author, but I think this paper really demonstrates the amazing power of retrospective multi-species systems modelling to provide insight into phenomena that are impossible to test empirically – i.e., questions of prehistoric (and in some cases, even data-poor historic) ecological change. The megafauna die-off controversy is one we’ve covered before here on ConservationBytes.com, and this is a related issue with respect to a charismatic extinction in Australia’s recent history – the loss of the Tasmanian thylacine (‘tiger’, ‘wolf’ or whatever inappropriate eutherian epithet one unfortunately chooses to apply). Read the rest of this entry »
The Philippine bare-backed fruit bat (Dobsonia chapmani; body size = < 220 mm, < 150 g; IUCN status: ‘Critically Endangered A2cd’) is endemic to lowland rain forests [top habitat image] from Negros and Cebu islands. This species of flying fox had been missing from the 1970s and was declared extinct in 2002 (34). In May 2003, five specimens [one shown in the picture above] were trapped in night nets in the Calatong forest (Negros Island), a ~ 1,000-ha fragment of secondary rain forest and agricultural lands [bottom habitat image] (35). The species is reliant on fruit-bearing vegetation and caves for feeding and roosting, respectively. As with many other Philippine bats, it suffers from habitat degradation and hunting. The family Pteropodidae comprises > 150 species. Despite their Draculian look, they all feed on fruits and nectar, and act as important plant pollinators (36), as well as disease vectors such as Ebola virus (37). Flying foxes are distributed in the tropics and subtropics from the Eastern Mediterranean, through the Arabian Peninsula, Asia, Australia, and many islands of the Indian Ocean. Photos courtesy of Ely L. Alcala.
Jared Diamond(1) coined the expression ‘evil quartet’ for the four main human causes of species extinctions: habitat loss/fragmentation, overkill, introduced species and extinction chains [with climate change and extinction synergies (2), the updated expression would be ‘evil sextet”]. However, one third of ‘extinct’ mammal species has been ‘found’ again. Recent studies reveal that the probability of rediscovery depends on the cause of extinction.
Arriving in a city to search for an old friend, I would first look in the suburb where he lived, the pub where we enjoyed a drink and some music, or the park where we used to play football. But if my friend was an outlaw, or had recently gone through a traumatic experience, my chances of finding him at his favourite spots would shrink.
If, instead of a friend, we are searching for the last survivors of an extinct-declared species, surveys also tend to take place in the habitat in which the species was previously found. Such a strategy rests on the classical hypothesis that, given the spatial distribution of a species, its gradual decline must occur from the periphery to the core of its distribution (‘range collapse’) where, in theory, the habitat should be of better quality and the number of individuals higher (3). In contrast, recent work supports that the trajectory of demise of threatened vertebrates progresses from the core to the periphery (‘range eclipse’) (4), because many perturbations make their way as a progressive wave, e.g, fire, logging or urbanisation.
Diana Fisher (5) supports the ‘range eclipse’ hypothesis for ‘extinct’ mammals which have been rediscovered. She quantifies that 60% of the new records are made from peripheral habitats, mainly when the principal cause of extirpation is habitat loss. Not only that, on average species are rediscovered at altitudes 35 % higher than historical records, and only in 5 % of the cases at the locality where it had been last seen.
Get Conservation Bytes by Email Enter your email address to receive new posts on conservation, biodiversity, and environmental science directly in your inbox. No need to visit the website regularly, and you can unsubscribe at any time.
Hulton Archive / Getty Images We warn readers that the content of this study is confronting and may be distressing. In April 1789, the first smallpox epidemic among Aboriginal people in the Sydney region began, just 16 months after Europeans arrived on the First Fleet. Little is known of the true scale of the epidemic,…
Just over two years ago I reported the ‘likely’ eradication of feral pigs from Australia’s third-largest (4,405 km2) island — Kangaroo Island. I indicated ‘likely’ because the program still required the proof-of-eradication phase to be completed before an official declaration could be made. Yesterday I had the immense honour to take part in the official…
Have you ever done any research that relied to any degree on Indigenous Knowledges? How did you cite those Knowledges, if at all? It’s probably time we rethink how we engage with Indigenous Knowledge systems. In a new article published in BioScience, we — a large group of Indigenous and non-Indigenous scholars in Australia —…





 We’ve just
We’ve just 







