A few weeks ago we published a paper that was in the works for a long time, so long in fact that one of my co-authors died before it was published online.
Paul Ehrlich died on 13 March 2026 at the ripe old age of 93, exactly two weeks before our article appeared online. Paul had a good innings no doubt, but I wish he had survived long enough to see what might very well be his last co-authored paper.
I first met Paul back in the mid-2000s during a trip through San Francisco. I had organised to come chat with Professor Gretchen Daily at Stanford, and Paul came along for lunch. I remember vividly how we clicked almost immediately.
We clicked so well in fact, that we wrote a book together, co-authored several high-impact papers (e.g., ‘ghastly future‘), spent a month in Bellagio as Rockefeller Foundation writing residents, participated in various public and parliamentary presentations, and generally just got on like a house on fire. Paul and his wife Ann became like family, so much so that they were de facto grandparents to my daughter who grew up with them in near-annual contact.
This post isn’t about Paul per se, but I cannot ignore the profound influence Paul had on my career, my personality, and my life view. I miss him. I am therefore dedicating this paper and post to his memory. So long, and thanks for all the fish.
Nothing like writing about human population to get the punters engaged.
We show empirically that the Earth has already exceeded its ability to support the global human population sustainably, with dire implications for increasing pressure on food security, climate stability, and human wellbeing. However, slowing population growth and raising global awareness could still offer us some hope.
Our study shows that humans have pushed well beyond the planet’s long-term carrying capacity and that continued growth under current patterns of consumption will intensify environmental and social challenges for communities worldwide.
Using animals as sport symbols reflects the integration of biodiversity into cultural identity and the transmission of collective values. This raises the possibility that the economic muscle of the sport industry could translate its symbolic capital into tangible commitments to biodiversity conservation.
Those who have had the privilege of travelling in remote areas might have come across an unexpected scene: a football pitch in the middle of the Amazon rain forest or on the slopes of the Andes, a basketball court on the side of a Buddhist temple, or an ice hockey rink on a snow-ploughed lake in remote northern Canada.
Sport is a global industry that generates identity, belonging, education, and shared emotions for both athletes and their avid spectators (1). Sporting affinities now rival the sense of nationhood once shared by citizens during warfare (2).
Now in our heavily monetised world, sport clubs rely on their fans through tickets and merchandising, and indirectly through television rights and advertising. In this both emotional and commercial relationship, expressions such as being true to the badge reflect the central role of corporate symbols in building bonds between a club and its supporters (3).
Sport club logos with animal iconography. Top row: examples of the grey wolf (Canis lupus) in Neftekhimik Nizhnekamsk (ice hockey, Russia), Warrington Wolves (rugby, England), Wolverhampton Wanderers (football, England), and Roma (football, Italy). Second row: bald eagle (Haliaeetus leucocephalus) in Essex Eagles (cricket, England), Adler Mannheim (handball, Germany), and Philadelphia Eagles (American football, USA). Third row: Free State Cheetahs (represented by the cheetah Acinonyx jubatus; rugby, South Africa), Toronto Blue Jays (blue jay Cyanocitta cristata; baseball, Canada), Memphis Grizzlies (grizzly bear Ursus arctos horribilis; basketball, USA) and Hisamitsu Springs (Japanese white-eye Zosterops japonicas; volleyball, Japan). Fourth row: six Spanish football clubs. Clubs featuring wolves and eagles are often associated with the symbolic qualities of these species (e.g., intelligence, prowess, fealty, bravery, strength). In football, animals reflect represent the history and heraldry of cities and regions, as seen in the crests of in Atlético de Madrid (brown bear Ursus arctos), AS Roma and Athletic Club (wolf), Cultural Deportiva Leonesa and Atlético Osasuna (lion Panthera leo), CD Castellón (raptor) and Levante UD (bat). Photos: Gary Kramer (wolf) and Andy Morffew (eagle).
In professional sport such as football, clubs increasingly function as brands (4) where even traditional logos are modified to enhance a team’s commercial value and strengthen audience loyalty (5). In this process, biodiversity becomes relevant because the iconography of many sport organisations incorporates representations of plants and animals.
Sport fauna
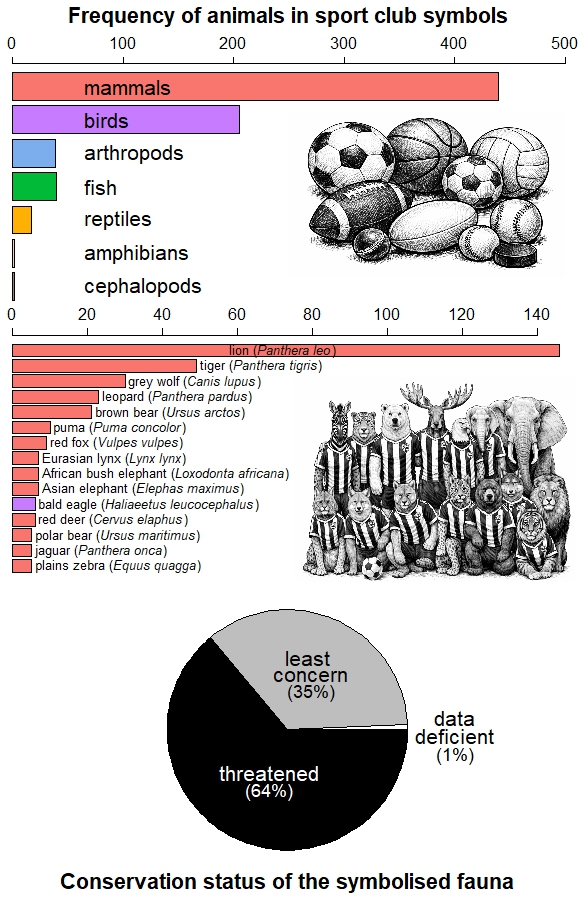
To quantify this phenomenon, Ugo Arbieu and collaborators analysed the presence of animals in club names, crests, and fan nicknames among 10 professional team sports across 50 countries (6). They found that 727 teams use 161 different animal species in their corporate imagery. Football and basketball lead in the number of species represented due to the large number of clubs worldwide, but American football, rugby and baseball display greater symbolic fauna diversity per club. Mammals and birds are the most common, particularly carnivores and raptors.
Animal symbols in club iconography (names, logos, fan nicknames) for the sports with the largest audiences (6): basketball, handball, baseball, cricket, football, American football, ice hockey and volleyball. The sample excludes 106 teams that use domesticated species as identity symbols, includes 163 men’s leagues and 67 women’s leagues, respectively, and the animal species depicted in the emblems of 48 teams could be identified. Horizontal bars above show the most represented animal groups (top panel), and the 15 species most frequently featured (middle panel). Bottom panel: percentage of symbols according to the IUCN’s conservation status of the species, where ‘threatened’ includes the categories Near Threatened, Endangered and Extinct. The trend indicates that sport clubs prefer to identify themselves with large mammal species that are threatened.
This pattern is not coincidental, for it reflects the historical bias of science and conservation towards large, charismatic vertebrates (7), but also the uneven availability of biological information and our social preferences for certain species (8). These preferences are even reflected in the animal emojis we share regularly on social media (9).
Arbieu’s study also revealed that clubs tend to favour images of threatened fauna (6), possibly due to their higher symbolic impact and media visibility (10). Moreover, although clubs in Europe and the Americas more often depict exotic animals, native species dominate in Africa, Asia, and Oceania (6). This suggests that the choice of an animal as an emblem is the product of not only aesthetic or symbolic criteria, but also of cultural roots and the historical relationship of societies with their local fauna.
Those of us living with cats share our homes with an ancestral predator, one adapted for hunting and the frequent, exclusive consumption of meat. These instincts become fully activated outside the domestic environment, where cats pose a global threat to wildlife.
Pets are family. We celebrate their arrival with the same joy as a grand homecoming, and their absence leaves a grief as deep as losing a loved one. In bonding with cats and dogs, we often attribute human abilities and emotions to them.
But beyond this affection, domestic animals still carry the instincts and genetic legacy of their wild ancestors(1, 2). My cats — Caruso, Muesli, and Plata — have been calm and loving, but they have always enjoyed a real hunt (3). When a moth comes in through a window, they seem possessed: their mouths chitter and make clicking sounds, they leap from one piece of furniture to another, and their heads snap sharply between the insect’s position and other points in the room, calculating the best spot from which to pounce on their prey. That is why when they become feral, cats and dogs integrate into food chains like any other species: they compete for ecosystem resources, hunt and are hunted, and hybridise and exchange diseases with other carnivores (4, 5).
Top: cat eating an Eurasian blue tit (Cyanistes caeruleus), a common visitor to home gardens in Nijmegen (Netherlands). Bottom, domestic cat after hunting an Eastern cottontail rabbit (Sylvilagus floridanus) in a residential neighbourhood of Stratford (Connecticut, USA). Photos courtesy of Jelger Herder (Nijmegen) and Scott Kruitbosch (Stratford). Scott is a photographer and conservationist. Near sunset on 30/09/2020, while intently observing local wildlife, he witnessed a neighbourhood cat sneak up from behind on a cottontail feeding in open grass and grab it. For years, Scott has had extremely negative interactions, both in person and online, with local residents over these issues. These exchanges have revealed that many people show little concern for wildlife or for the dangers their outdoor cats face, and believe that their cats would not, or could not, harm wildlife.
Domestic cats are highly skilled hunters, and their predatory interactions with a wide range of prey are widely documented in social media and documentaries. Some examples include cats catching: bats and birds on the wing, butterflies, chipmunks, dragons, fishes, grasshoppers, frogs, lizards, mice, owls, rabbits, seagulls, snakes, squirrels, and wallabies. See an award-winning photo depicting wildlife with fatal injuries caused by cats recorded in 2019 at a single animal hospital in the USA, and a video showing domestic cats mimicking bird calls and some cat owners explaining that their pets reject commercial cat food after experiencing the thrill of hunting real prey. The documentary Secret Life of Cats contextualises the ecological challenges posed by free-roaming cats.
Coral reefs are much more than just a pretty place to visit. They are among the world’s richest ecosystems, hosting about a third of all marine species.
These reefs also directly benefit more than a billion people, providing livelihoods and food security, as well as protection from storms and coastal erosion.
Without coral reefs, the world would be a much poorer place. So when corals die or become damaged, many people try to restore them. But the enormity of the task is growing as the climate keeps warming.
In our new research, we examined the full extent of existing coral restoration projects worldwide. We looked at what drives their success or failure, and how much it would actually cost to restore what’s already been lost. Restoring the reefs we’ve already lost around the world could cost up to A$26 trillion.
Bleached Acropora corals in the Maldives.Davide Seveso/University of Milan
When sea temperatures climb above the seasonal average for sustained periods, corals can become bleached. They lose colour as they expel their symbiotic algae when stressed, revealing the white skeleton underneath. Severe bleaching can kill coral.
Human overpopulation is often depicted in the media in one of two ways: as either a catastrophic disaster or an overly-exaggerated concern. Yet the data understood by scientists and researchers is clear. So what is the actual state of our overshoot, and, despite our growing numbers, are we already seeing the signs that the sixth mass extinction is underway?
In a recent episode of The Great Simplification podcast, Nate Hagens was joined by global ecologist Corey Bradshaw to discuss his recent research on the rapid decline in biodiversity, how population and demographics will change in the coming decades, and what both of these will mean for complex global economies currently reliant on a stable environment.
Australia is home to about one in 12 of the world’s species of animals, birds, plants and insects – between 600,000 and 700,000 species. More than 80% of Australian plants and mammals and just under 50% of our birds are found nowhere else.
But habitat destruction, climate change, and invasive species are wreaking havoc on Earth’s rich biodiversity, and Australia is no exception.
More and more species stand on the edge of oblivion. That’s just the ones we know enough about to list formally as threatened. Many more are in trouble, especially in the oceans. Change is the new constant. As the world heats up and ecosystems warp, new combinations of species can emerge without an evolutionary connection, creating novel communities.
It is still possible to stop species from dying out. But it will take an unprecedented effort.
The vulnerable southern bell (growling grass) frog (Litoria raniformis). Rupert Mathwin/Flinders University
In light of new genetic research on the identity of ‘wild dogs’ and dingoes across Australia, the undersigned wish to express concern with current South Australia Government policy regarding the management and conservation of dingoes. Advanced DNA research on dingoes has demonstrated that dingo-dog hybridisation is much less common than thought, that most DNA tested dingoes had little domestic dog ancestry and that previous DNA testing incorrectly identified many dingoes as hybrids (Cairns et al. 2023). We have serious concerns about the threat current South Australian public policy poses to the survival of the ‘Big Desert’ dingo population found in Ngarkat Conservation Park and surrounding areas.
We urge the South Australian Government to:
Revoke the requirement that all landholders follow minimum baiting standards, including organic producers or those not experiencing stock predation. Specifically
Dingoes in Ngarkat Conservation park (Region 4) should not be destroyed or subjected to ground baiting and trapping every 3 months. The Ngarkat dingo population is a unique and isolated lineage of dingo that is threatened by inbreeding and low genetic diversity. Dingoes are a native species and all native species should be protected inside national parks and conservation areas.
Landholders should not be required to carry out ground baiting on land if there is no livestock predation occurring. Furthermore, landholders should be supported to adopt non-lethal tools and strategies to mitigate the risk of livestock predation including the use of livestock guardian animals, which are generally incompatible with ground and aerial 1080 baiting.
Revoke permission for aerial baiting of dingoes (incorrectly called “wild dogs”) in all Natural Resource Management regions – including within national parks. Native animals should be protected in national parks and conservation areas.
Cease the use of inappropriate and misleading language to label dingoes as “wild dogs”. Continued use of the term “wild dogs” is not culturally respectful to First Nations peoples and is not evidence-based.
Proactively engage with First Nations peoples regarding the management of culturally significant species like dingoes. For example, the Wotjobaluk nation should be included in consultation regarding the management of dingoes in Ngarkat Conservation Park.
Changes in South Australia public policy are justified based on genetic research by Cairns et al. (2023) that overturns previous misconceptions about the genetic status of dingoes. It demonstrates:
Most “wild dogs” DNA tested in arid and remote parts of Australia were dingoes with no evidence of dog ancestry. There is strong evidence that dingo-dog hybridisation is uncommon, with firstcross dingo-dog hybrids and feral dogs rarely being observed in the wild. In Ngarkat Conservation park none of DNA tested animals had evidence of domestic dog ancestry, all were ‘pure’ dingoes.
Previous DNA testing methods misidentified pure dingoes as being mixed. All previous genetic surveys of wild dingo populations used a limited 23-marker DNA test. This is the method currently used by NSW Department of Primary Industries, which DNA tests samples from NSW Local Land Services, National Parks and Wildlife Service, and other state government agencies. Comparisons of DNA testing methods find that the 23-marker DNA test frequently misidentified animals as dingo-dog hybrids. Existing knowledge of dingo ancestry across South Australia, particularly from Ngarkat Conservation park is incorrect; policy needs to be based on updated genetic surveys.
There are multiple dingo populations in Australia. High-density genomic data identified more than four wild dingo populations in Australia. In South Australia there are at least two dingo populations present: West and Big Desert. The West dingo population was observed in northern South Australia, but also extends south of the dingo fence. The Big Desert population extends from Ngarkat Conservation park in South Australia into the Big Desert and Wyperfield region of Victoria.
The Ngarkat Dingo population is threatened by low genetic variability. Preliminary evidence from high density genomic testing of dingoes in Ngarkat Conservation park and extending into western Victoria found evidence of limited genetic variability which is a serious conservation concern. Dingoes in Ngarkat and western Victoria had extremely low genetic variability and no evidence of gene flow with other dingo populations, demonstrating their effective isolation. This evidence suggests that the Ngarkat (and western Victorian) dingo population is threatened by inbreeding and genetic isolation. Continued culling of the Ngarkat dingo population will exacerbate the low genetic variability and threatens the persistence of this population.
But just how far-fetched is it? Could pathogens that were once common on Earth – but frozen for millennia in glaciers, ice caps and permafrost – emerge from the melting ice to lay waste to modern ecosystems? The potential is, in fact, quite real.
In 2014, a giant “zombie” Pithovirus sibericum virus was revived from 30,000-year-old Siberian permafrost.
And in 2016, an outbreak of anthrax (a disease caused by the bacterium Bacillus anthracis) in western Siberia was attributed to the rapid thawing of B. anthracis spores in permafrost. It killed thousands of reindeer and affected dozens of people.
Bacillus anthracis is a soil bacterium that causes anthrax. William A. Clark/USCDCP
More recently, scientists found remarkable genetic compatibility between viruses isolated from lake sediments in the high Arctic and potential living hosts.
Earth’s climate is warming at a spectacular rate, and up to four times faster in colder regions such as the Arctic. Estimates suggest we can expect four sextillion (4,000,000,000,000,000,000,000) microorganisms to be released from ice melt each year. This is about the same as the estimated number of stars in the universe.
However, despite the unfathomably large number of microorganisms being released from melting ice (including pathogens that can potentially infect modern species), no one has been able to estimate the risk this poses to modern ecosystems.
In a new study published today in the journal PLOS Computational Biology, we calculated the ecological risks posed by the release of unpredictable ancient viruses.
Have you ever watched a nature documentary and marvelled at the intricate dance of life unfolding on screen? From the smallest insect to the largest predator, every creature plays a role in the grand performance of our planet’s biosphere. But what happens when one of these performers disappears?
In this post, we delve into our recent article Estimating co-extinction risks in terrestrial ecosystems just published in Global Change Biology, in which we discuss the cascading effects of species loss and the risks of ‘co-extinction’.
But what does ‘co-extinction’ really mean?
Imagine an ecosystem as a giant web of interconnected species. Each thread represents a relationship between two species — for example, a bird that eats a certain type of insect, or a plant that relies on a specific species of bee for pollination. Now, what happens if one of these species in the pair disappears? The thread breaks and the remaining species loses an interaction. This could potentially lead to its co-extinction, which is essentially the domino effect of multiple species losses in an ecosystem.
A famous example of this effect can be seen with the invasion of the cane toad (Rhinella marina) across mainland Australia, which have caused trophic cascades and species compositional changes in these communities.
The direct extinction of one species, caused by effects such as global warming for example, has the potential to cause other species also to become extinct indirectly.
Following my annual tradition, I present the retrospective list of the ‘top’ 20 influential papers of 2022 as assessed by experts in Faculty Opinions(formerly known as F1000). These are in no particular order. See previous years’ lists here: 2021, 2020, 2019, 2018, 2017, 2016, 2015, 2014, and 2013.
Climate change is one of the main drivers of species loss globally. We know more plants and animals will die as heatwaves, bushfires, droughts and other natural disasters worsen.
But to date, science has vastly underestimated the true toll climate change and habitat destruction will have on biodiversity. That’s because it has largely neglected to consider the extent of “co-extinctions”: when species go extinct because other species on which they depend die out.
Our new research shows 10% of land animals could disappear from particular geographic areas by 2050, and almost 30% by 2100. This is more than double previous predictions. It means children born today who live to their 70s will witness literally thousands of animals disappear in their lifetime, from lizards and frogs to iconic mammals such as elephants and koalas.
But if we manage to dramatically reduce carbon emissions globally, we could save thousands of species from local extinction this century alone.
Ravages of drought will only worsen in coming decades. CJA Bradshaw
An extinction crisis unfolding
Every species depends on others in some way. So when a species dies out, the repercussions can ripple through an ecosystem.
For example, consider what happens when a species goes extinct due to a disturbance such as habitat loss. This is known as a “primary” extinction. It can then mean a predator loses its prey, a parasite loses its host or a flowering plant loses its pollinators.
A real-life example of a co-extinction that could occur soon is the potential loss of the critically endangered mountain pygmy possum (Burramys parvus) in Australia. Drought, habitat loss, and other pressures have caused the rapid decline of its primary prey, the bogong moth (Agrotis infusa).
The conservation, environment, and sustainability literature is rife with the term ‘collapse’, applied to concepts as diverse as species extinction to the complete breakdown of civilisation. I have also struggled with its various meanings and implications, so I’m going to attempt to provide some clarity on collapse for my own and hopefully some others’ benefit.
From a strictly ecological perspective, ‘collapse’ could be described in the following (paraphrased) ways:
abrupt transition of one ecosystem state to another, usually invoking the idea that something has declined in the process (species richness, beta diversity, functional diversity, trophic network connectance, trait volume, production, etc.);
But there is still nor formal definition of ‘collapse’ in ecology, as identified by several researchers (Keith et al. 2013; Boitani et al. 2015; Keith et al. 2015; Sato and Lindenmayer 2017; Bland et al. 2018). While this oversight has been discussed extensively with respect to quantifying changes, I can find nothing in the literature that attempts a generalisable definition of what collapse should mean. Perhaps this is because it is not possible to identify a definition that is sufficiently generalisable, something that Boitani et al. (2015) described with this statement:
“The definition of collapse is so vague that in practice it will be possible (and often necessary) to define collapse separately for each ecosystem, using a variety of attributes and threshold values
Despite all the work that has occurred since then, I fear we haven’t moved much beyond that conclusion.
Hell, cutting down the trees in the bush block next to my property constitutes a wholesale ‘collapse’ of the microcommunity of species using that patch of bush. An asteroid hitting the Earth and causing a mass extinction is also collapse. And everything in-between.
But at least ecologists have made some attempts to define and quantify collapse, even if an acceptable definition has not been forthcoming. The sustainability and broader environment literature has not even done that.
In a newly announced partnership with Texas biotech company Colossal Biosciences, Australian researchers are hoping their dream to bring back the extinct thylacine is a “giant leap” closer to fruition.
Scientists at University of Melbourne’s TIGRR Lab (Thylacine Integrated Genetic Restoration Research) believe the new partnership, which brings Colossal’s expertise in CRISPR gene editing on board, could result in the first baby thylacine within a decade.
The genetic engineering firm made headlines in 2021 with the announcement of an ambitious plan to bring back something akin to the woolly mammoth, by producing elephant-mammoth hybrids or “mammophants”.
But de-extinction, as this type of research is known, is a highly controversial field. It’s often criticised for attempts at “playing God” or drawing attention away from the conservation of living species. So, should we bring back the thylacine? We asked five experts.
Now that Clarivate, Google, and Scopus have recently published their respective journal citation scores for 2021, I can now present — for the 14th year running on ConvervationBytes.com — the 2021 conservation/ecology/sustainability journal ranks based on my journal-ranking method.
Like last year, I’ve added a few journals. I’ve also included in the ranking the Journal Citation Indicator (JCI) in addition to the Journal Impact Factor and Immediacy Index from Clarivate ISI, and the CiteScore (CS) in addition to the Source-Normalised Impact Per Paper (SNIP) and SCImago Journal Rank (SJR) from Scopus.
I therefore present the new 2021 ranks for: (i) 106 ecology, conservation and multidisciplinary journals, (ii) 27 open-access (i.e., you have to pay) journals from the previous category, (iii) 64 ‘ecology’ journals, (iv) 32 ‘conservation’ journals, (v) 43 ‘sustainability’ journals (with general and energy-focussed journals included), and (vi) 21 ‘marine & freshwater’ journals.
Remember not to take much notice if a journal boasts about how its Impact Factor has increased this year, because these tend to increase over time anyway What’s important is a journal’s relative (to other journals) rank.
Nearly a decade ago (my how time flies*), I wrote a post about the guaranteed failure of government policies purporting no-extinction targets within their environmental plans. I was referring to the State of South Australia’s (then) official policy of no future extinctions.
In summary, zero- (or no-) extinction targets at best demonstrate a deep naïvety of how ecology works, and at worst, waste a lot of resources on interventions doomed to fail.
4. Climate change will also guarantee additional (perhaps even most) future extinctions irrespective of Australian policies.
I argued that no-extinction policies are therefore disingenuous to the public in the extreme because they sets false expectations, engender disillusionment after inevitable failure, and ignores the concept of triage — putting our environment-restoration resources toward the species/systems with the best chance of surviving (uniqueness notwithstanding).
Bees are essential for pollination, but their critical function can be perturbed by pesticides. The detrimental effects of those chemicals accumulate through a bee’s life, and become stronger if females cannot collect pollen from wildflowers.
Our childhood experiences partly determine our health, personality, and lifestyle when we are adults, and our experiences accumulate over time. Accumulation also occurs in any living being and can explain why some populations and species adapt to their environments better than others.
Migratory birds are a clear example. Thousands can travel to their breeding grounds after wintering elsewhere, and those coming from regions laden with resources (e.g., food, shelter, water) will have a greater reproductive success than those that migrated from resource-poor regions (1). In ecology, these ‘carry-over’ effects can take place between seasons, but also across the different phases of the life cycle of a plant or animal (2).
From larvae to adults
Clara Stuligross and Neal Williams have studied the carry-over effect of pesticides on the blue orchard beeOsmia lignaria in California (3). Instead of the typical hives constructed by the honey bee (Apis mellifera), solitary blue orchard bees make lines of brood cells with mud partitions, glued into holes and crevices of branches and trunks from fallen trees (see videos here, here, & here).
Carnivores are essential components of trophic webs, and ecosystem functions crumble with their loss. Novel data show the connection between calcareous reefs and sea otters under climate change.
Trophic cascade on the Aleutian Islands (Alaska, USA) linking sea otters (Enhydra lutris) with sea urchins (Strongylocentrotus polyacanthus) and calcareous reefs (Clathromorphum nereostratum). With males weighting up to 50 kg, sea otters have been IUCN-catalogued as Endangered since 2000. The top photo shows a male in a typical, belly-up floating position. The bottom photo shows live (pinkish) and dead (whitish) tissue on the reef surface as a result of grazing of sea urchins at a depth of 10 m. Sea otters are mesopredators, typically foraging on small prey like sea urchins, but their historical decline due to overhunting unleashed the proliferation of the echinoderms. At the same time, acidification and sea-water warming have softened the skeleton of the reefs, allowing for deeper grazing by sea urchins that eliminate the growth layer of living tissue that give the reefs their pinkish hue. Large extents of dead reefs stop fixing the excess in carbonic acid, whose carbon atoms sea water sequesters from the atmosphere enriched in carbon by our burning of fossil fuels. Photos courtesy of Joe Tomoleoni taken in Moss Landing – California, USA (otter), and on the Near Islands – Aleutian Archipelago, Alaska (reef).
For most, the decisions made by people we have never met affect our daily lives. Other species experience the same phenomenon because they are linked to one another through a trophic cascade.
A trophic cascade occurs when a predator limits the abundance or behaviour of its prey, in turn affecting the survival of a third species in lower trophic levels that have nothing directly to do with the predator in question (1).
Sea otters (Enhydra lutris) represent a text-book example of a trophic cascade. These mustelids (see video footage here and here) hunt and control the populations of sea urchins (Strongylocentrotus polyacanthus), hence favouring kelp forests — the fronds of which are eaten by the sea urchins.
Removing the predator from the equation should lead to more sea urchins and less kelp, and this chain of events is exactly what happened along the coasts of the North Pacific (2, 3). The historical distribution of sea otters once ranged from Japan to Baja California through the Aleutian Islands (see NASA’s photo from space, and documentary on the island of Unimak), a sub-Arctic, arc-shaped archipelago including > 300 islands between Alaska (USA) and the Kamchatka Peninsula (Russia), extending ~ 2000 kilometres, and having a land area of ~ 18,000 km2.
But the fur trade during the 18th and 19th centuries brought the species to the brink of extinction, down to < 2000 surviving individuals (4). Without otters, sea urchins boomed and deforested kelp ecosystems during the 20th Century (5). Now we also know that this trophic cascade has climate-related implications in other parts of the marine ecosystem.
Underwater bites
Doug Rasher and collaborators have studied the phenomenon on the Aleutian Islands (6). The seabed of this archipelago is a mix of sandy beds, kelp forests, and calcareous reefs made up of calcium and magnesium carbonates fixed by the red algae Clathromorphum nereostratum. These reefs have grown at a rate of 3 cm annually for centuries as the fine film of living tissue covering the reef takes the carbonates from the seawater (7).
Last week, researchers at the University of Melbourne announced that thylacines or Tasmanian tigers, the Australian marsupial predators extinct since the 1930s, could one day be ushered back to life.
The thylacine (Thylacinus cynocephalus), also known as the ‘Tasmanian tiger’ (it was neither Tasmanian, because it was once common in mainland Australia, nor was it related to the tiger), went extinct in Tasmania in the 1930s from persecution by farmers and habitat loss. Art by Eleanor (Nellie) Pease, University of Queensland. Centre of Excellence for Australian Biodiversity and Heritage
Advances in mapping the genome of the thylacine and its living relative the numbat have made the prospect of re-animating the species seem real. As an ecologist, I would personally relish the opportunity to see a living specimen.
The announcement led to some overhyped headlines about the imminent resurrection of the species. But the idea of “de-extinction” faces a variety of technical, ethical and ecological challenges. Critics (like myself) argue it diverts attention and resources from the urgent and achievable task of preventing still-living species from becoming extinct.
The rebirth of the bucardo
The idea of de-extinction goes back at least to the the creation of the San Diego Frozen Zoo in the early 1970s. This project aimed to freeze blood, DNA, tissue, cells, eggs and sperm from exotic and endangered species in the hope of one day recreating them.
The notion gained broad public attention with the first of the Jurassic Park films in 1993. The famous cloning of Dolly the sheep reported in 1996 created a sense that the necessary know-how wasn’t too far off.
The next technological leap came in 2008, with the cloning of a dead mouse that had been frozen at –20℃ for 16 years. If frozen individuals could be cloned, re-animation of a whole species seemed possible.
After this achievement, de-extinction began to look like a potential way to tackle the modern global extinction crisis.
No matter most people’s best intentions, poaching of species in Sub-Saharan Africa for horn and ivory continues unabated. Despite decades of policies, restrictions, interventions, protections, and incentives, many species of elephant and rhino are still hurtling toward extinction primarily because of poaching.
Clearly, we’re doing something heinously wrong.
Collectively, we have to take a long, hard look in the conservation mirror and ask ourselves some difficult questions. Why haven’t we been able to put any real dent in the illegal trade of poached elephant ivory and rhino horn? How many millions (billions?) of dollars have we spent seemingly to little avail? Why haven’t trade bans and intensive security measures done the trick?
The reasons are many, but they boil down to two main culprits:
neo-colonial sentiments driven by the best intentions of mainly overseas NGOs have inadvertently created the ideal conditions for the poaching economy — what we term poachernomics — to thrive by ensuring the continued restriction of legal supply of wildlife products; and
shutting off conservation areas to local people and directing the bulk of ecotourism profits away from source communities have maintained steady poaching incentives in the absence of other non-destructive livelihoods.
The logic of money contradicts the logic of species conservation and human health. As illegal trade has driven pangolins to near extinction, their hunting and market value has kept increasing ― even when we have known that they act as coronavirus reservoirs in the middle of the Covid-19 pandemic.
Sunda pangolin (Manis javanica) in a monsoon forest (Sumba Island, Indonesia). With adult weights up to 10 kg and body lengths around half a metre, these animals are mostly solitary and nocturnal, feed on ants and termites, and love tree climbing using bark hollows to shelter and give birth to singletons. The species occurs across mainland and islands of South East Asia, and became ‘Endangered’ in 2008 and ‘Critically Endangered’ in 2014, following a 80% decline in the last 20 years due to hunting and poaching. It has been the most heavily trafficked Asian species, and the IUCN’s assessment states: “… the incentives for harvesting and illegally trading in the species are universally high based on the high financial value of pangolin parts and derivatives”. Captive breeding is unlikely to deter wild collection because (among other reasons) farming costs are high (more so on a large scale) and, even if the species could be traded legally, wild versus farmed pangolin products and individuals are difficult to distinguish (23). Photo courtesy of Michael Pitts
Urbanites are attracted to exotic species, materials, and places. Our purchasing power seems to give us the right to buy any ‘object’ that we can pay for, no matter how exotic the object might be. In such a capitalist rationale, it is no surprise that > 150 thousand illegal cargos with wild animals and plants have been confiscated in 149 countries over the last two decades, moving some 6000 species from one place of the planet to another (1).
Social networks show people interacting with all kinds of fauna, creating the illusion that any animal can become a pet (2). And there’s a multi-$billion market of wildlife for a diverse array of uses including collecting, food, ornamentation, leisure, clothing and medicine (3-5). The paradox is that the rarer a species is, the higher its market value runs and the more lucrative selling it turns out to be, leading to more exploitation and rocketing extinction risk (6).
Hulton Archive / Getty Images We warn readers that the content of this study is confronting and may be distressing. In April 1789, the first smallpox epidemic among Aboriginal people in the Sydney region began, just 16 months after Europeans arrived on the First Fleet. Little is known of the true scale of the epidemic,…
Just over two years ago I reported the ‘likely’ eradication of feral pigs from Australia’s third-largest (4,405 km2) island — Kangaroo Island. I indicated ‘likely’ because the program still required the proof-of-eradication phase to be completed before an official declaration could be made. Yesterday I had the immense honour to take part in the official…
Have you ever done any research that relied to any degree on Indigenous Knowledges? How did you cite those Knowledges, if at all? It’s probably time we rethink how we engage with Indigenous Knowledge systems. In a new article published in BioScience, we — a large group of Indigenous and non-Indigenous scholars in Australia —…