Below we italicise each question and the name of the person asking the question and cite a range of publications we deem relevant per question. For expanding on Katharyne Hayhoe’s views on climate change, see a sample of her public talks here and here, interviews here and here, and newspaper articles here and here. We love one of the titles of her newspaper articles “A thermometer is not liberal or conservative”. A spanish version of this article and interview has been published in the February 2021 issue of the magazine Quercus.
Climate change has some localised, short-term, specific benefits (9). One example is increased access to oil and gas resources in a melting Arctic (10). This temporarily profits oil and gas industries, provides some financial benefit to local communities in Greenland and Alaska short-term, and harms both them and everyone else in the long term. A book called Windfall by Mackenzie Funk describes who is “profiteering” from climate change, and how.
Overall, however, climate change already harms the majority of people today. The poor, the vulnerable, and the marginalized are affected first and foremost. Since the 1960s, for example, climate change has increased the gap between the richest and poorest countries in the world by as much as 25 per cent. In 2019, UN Special Rapporteur on extreme poverty and human rights, Philip Alston, warned that climate change “threatens to undo the last 50 years” of development, global health and poverty reduction.” (11)
And while the rich may be able to temporarily “buy their way out of rising heat and hunger”, as Alston put it, the truth is that we all live on this planet, no matter how wealthy and influential we are. The air we breathe, the water we drink, the food we eat and all the resources we use come from our shared home.
Climate change threatens the ability of our planet to support human civilisation as we know it. It is a threat multiplier, attacking our health, our economy, our resources and even our security. As climate change intensifies and economic markets crumble and refugee crises surge, even those who may temporarily benefit from a warmer world will be negatively impacted by these changes long-term.
That’s why it makes so much sense to take practical steps to limit carbon pollution now. Many of these actions also provide us with short-term benefits that can be quantified in economic terms: like energy savings through efficiency, cheaper electricity from renewables, more jobs, better public transportation, and even faster cars (like Tesla). Climate action also provides less tangible but arguably even more important benefits: cleaner air and water, better health, poverty reduction, and a host of other co-benefits that substantively move us towards meeting key UN Sustainable Development Goals.
To care about climate change, we don’t have to be a certain type of person or live in a certain place or vote a certain way: all we have to be is a human living on this planet, and we’re all that.







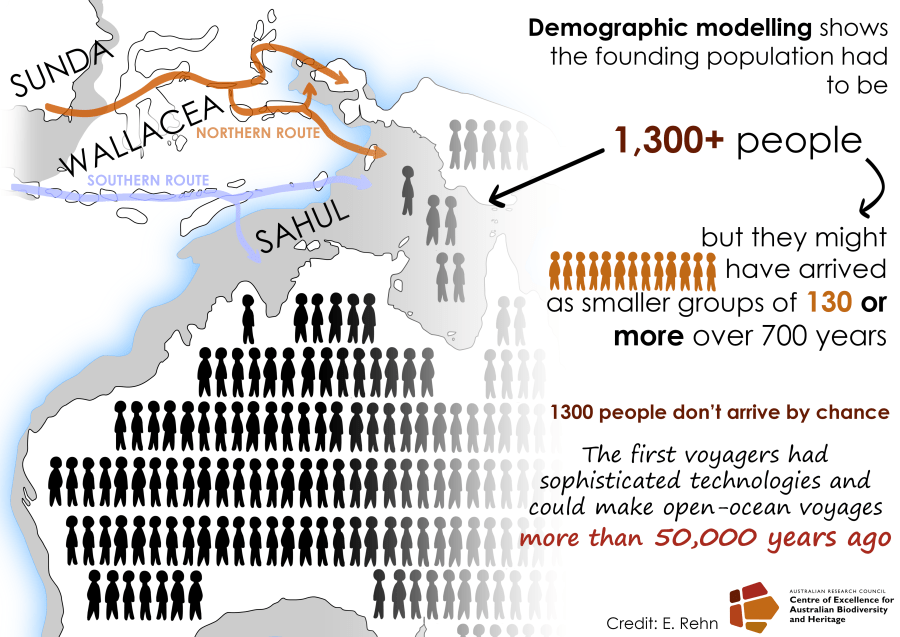

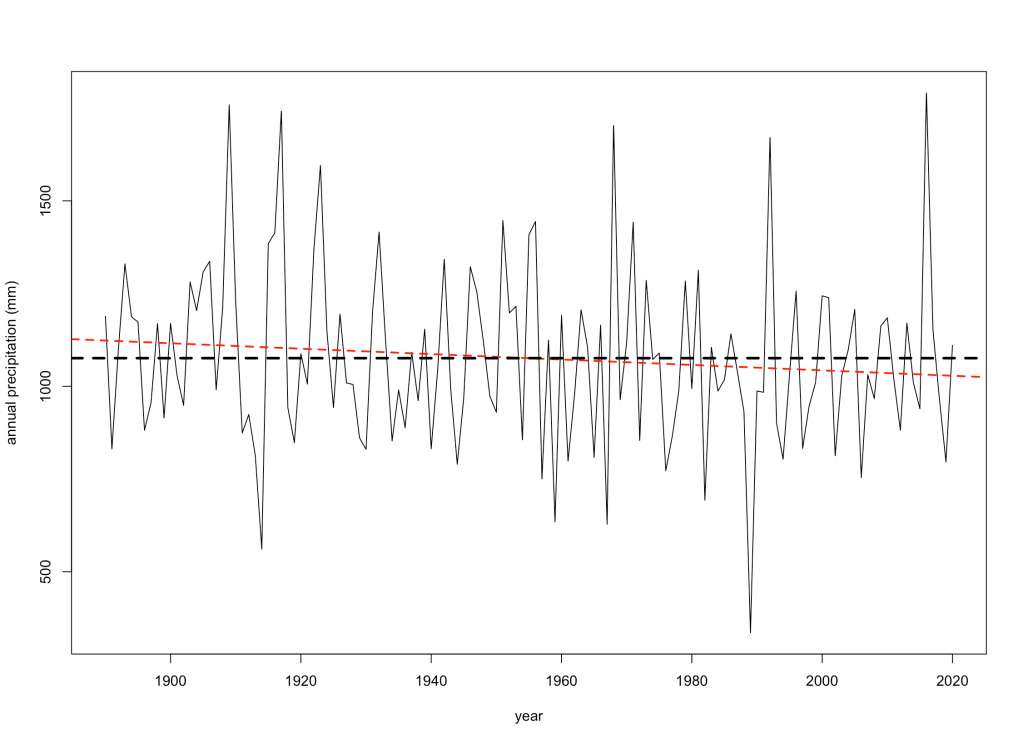











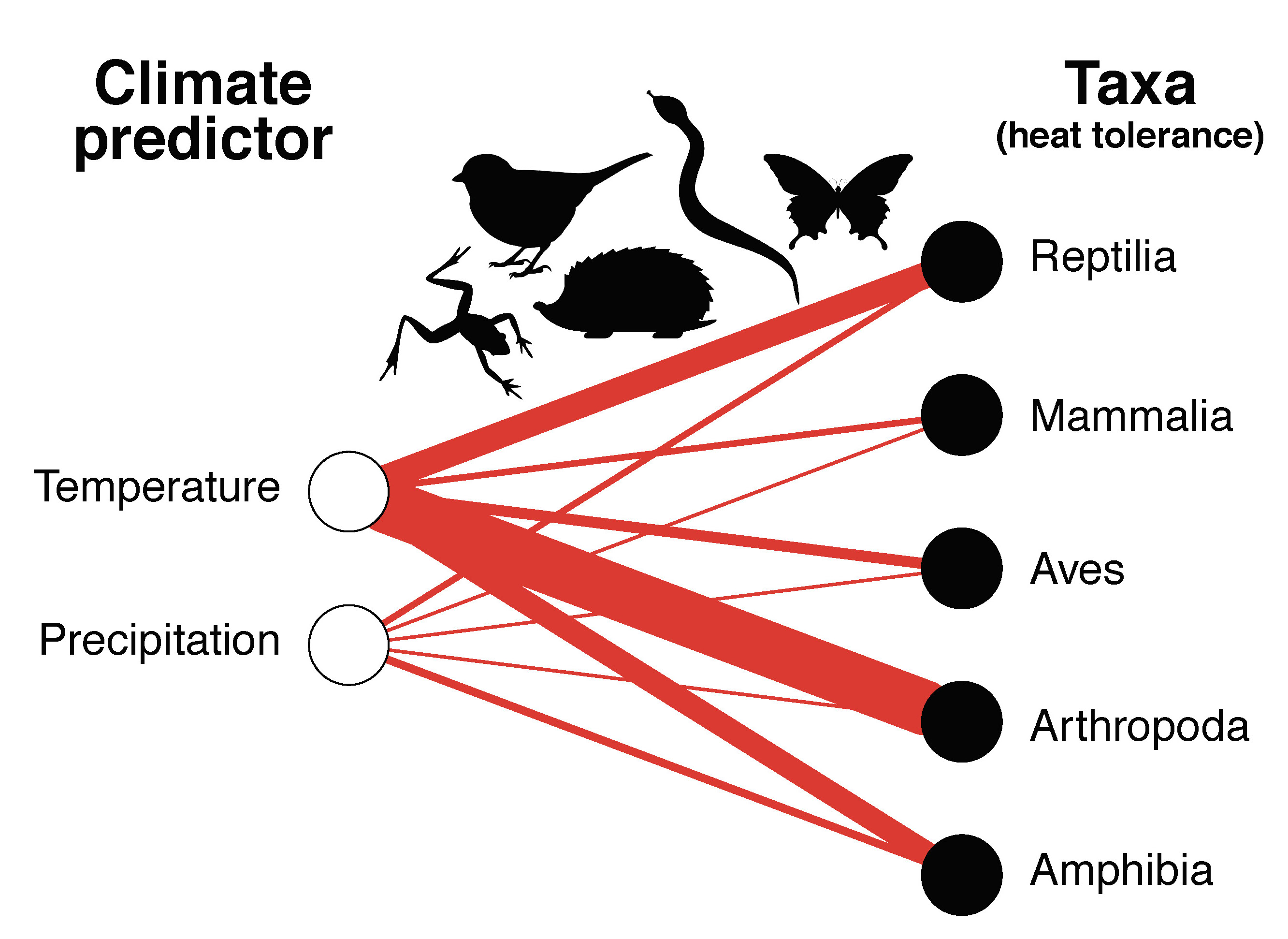
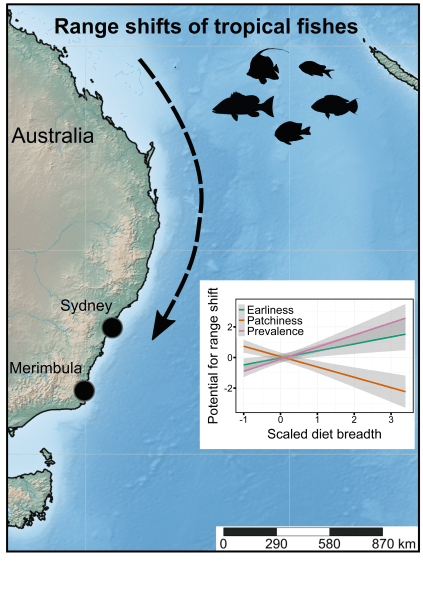 Ecologists often rely on measuring certain elements of a species’ characteristics, behaviour, or morphology to determine if these — what we call ‘traits’ — give them certain capacities to exploit their natural environments. While sometimes a bit arbitrarily defined, the traits that can be measured are many indeed, and sometimes they reveal rather interesting elements of a species’ resilience in the face of environmental change.
Ecologists often rely on measuring certain elements of a species’ characteristics, behaviour, or morphology to determine if these — what we call ‘traits’ — give them certain capacities to exploit their natural environments. While sometimes a bit arbitrarily defined, the traits that can be measured are many indeed, and sometimes they reveal rather interesting elements of a species’ resilience in the face of environmental change.





