Today’s post comes from Salvador Herrando-Pérez (who, incidentally, recently submitted his excellent PhD thesis).
—

Three species co-occurring in the Gulf of Mexico and involved in the trophic cascade examined by Myers et al. (8). [1] Black-tips (Carcharhinus limbatus) are pelagic sharks in warm and tropical waters worldwide; they reach < 3 m in length, 125 kg in weight, with a maximum longevity in the wild of ~ 12 years; a viviparous species, with females delivering up to 10 offspring per parturition. [2] The cownose ray (Rhinoptera bonasus) is a tropical species from the western Atlantic (USA to Brazil); up to 2 m wide, 50 kg in weight, and 18 years of age; gregarious, migratory and viviparous, with one single offspring per litter. [3] The bay scallop (Agropecten irradians) is a protandric (hermaphrodite) mollusc, with sperm being released a few days before the (> 1 million) eggs; commonly associated with seagrasses in the north-western Atlantic; shells can reach up to 10 cm and individuals live for < 2 years. In the photos, a black-tip angled in a bottom long-line off Alabama (USA), a school of cownose rays swimming along Fort Walton Beach (Florida, USA), and a bay scallop among fronds of turtle grass (Thalassia testudinum) off Hernando County (Florida, USA). Photos by Marcus Drymon, Dorothy Birch and Janessa Cobb, respectively.
The hips of John Travolta, the sword of Luke Skywalker, and the teeth of Jaws marked an era. I still get goose pimples with the movie soundtrack (bass, tuba, orchestra… silence) solemnizing each of the big shark’s attacks. The media and cinema have created the myth of man’s worst friend. This partly explains why shark fishing does not trigger the same societal rejection as the hunting of other colossuses such as whales or elephants. Some authors contend that we currently live in the sixth massive extinction event of planet Earth (1) 75 % of which is strongly driven by one species, humans, and characterized by the systematic disappearance of mega-animals in general (e.g., mammoths, Steller’s seacow), and predators in particular, e.g., sharks (2, 3).
The selective extirpation of apex predators, recently coined as ‘trophic downgrading’, is transforming habitat structure and species composition of many ecosystems worldwide (4). In the marine realm, over the last half a century, the main target of the world’s fisheries has turned from (oft-large body-sized) piscivorous to planctivorous fish and invertebrates, indicating that fishery fleets are exploiting a trophic level down to collapse, then harvesting the next lower trophic level (5-7).
Myers et al. (8) illustrate the problem with the fisheries of apex-predator sharks in the northeastern coast of the USA. Those Atlantic waters are rife with many species of shark (> 2 m), whose main prey are smaller chondrichthyans (skates, rays, catsharks, sharks), which in turn prey on bottom fishes and bivalves. Myers et al. (8) found that, over the last three decades, the abundance of seven species of large sharks declined by ~ 90 %, coinciding with the crash of a centenary fishery of bay scallops (Agropecten irradians). Conversely, the abundance of 12 smaller chondrichthyes increased dramatically over the same period of time. In particular, the cownose ray (Rhinoptera bonasus), the principal predator of bay scallops, might today exceed > 40 million individuals in some bays, and consume up to ~ 840,000 tonnes of scallops annually. The obvious hypothesis is that the reduction of apex sharks triggers the boom of small chondrichthyans, hence leading to the break-down of scallop stocks. Read the rest of this entry »

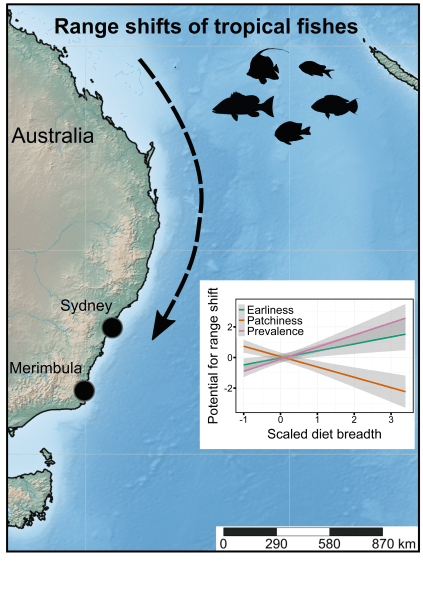 Ecologists often rely on measuring certain elements of a species’ characteristics, behaviour, or morphology to determine if these — what we call ‘traits’ — give them certain capacities to exploit their natural environments. While sometimes a bit arbitrarily defined, the traits that can be measured are many indeed, and sometimes they reveal rather interesting elements of a species’ resilience in the face of environmental change.
Ecologists often rely on measuring certain elements of a species’ characteristics, behaviour, or morphology to determine if these — what we call ‘traits’ — give them certain capacities to exploit their natural environments. While sometimes a bit arbitrarily defined, the traits that can be measured are many indeed, and sometimes they reveal rather interesting elements of a species’ resilience in the face of environmental change.