Coral reefs are much more than just a pretty place to visit. They are among the world’s richest ecosystems, hosting about a third of all marine species.
These reefs also directly benefit more than a billion people, providing livelihoods and food security, as well as protection from storms and coastal erosion.
Without coral reefs, the world would be a much poorer place. So when corals die or become damaged, many people try to restore them. But the enormity of the task is growing as the climate keeps warming.
In our new research, we examined the full extent of existing coral restoration projects worldwide. We looked at what drives their success or failure, and how much it would actually cost to restore what’s already been lost. Restoring the reefs we’ve already lost around the world could cost up to A$26 trillion.
Bleached Acropora corals in the Maldives.Davide Seveso/University of Milan
When sea temperatures climb above the seasonal average for sustained periods, corals can become bleached. They lose colour as they expel their symbiotic algae when stressed, revealing the white skeleton underneath. Severe bleaching can kill coral.
Night is the peak activity period for many animal species. In the Western Andes of Ecuador, the Chocó golden scarab flies between forest patches during the night, but urban lighting interferes with their paths and jeopardises populations already struggling to persist in fragmented native forests.
Urban development has created a network of illuminated infrastructure that allows our society to function day and night without interruption. It is no surprise that with so much artificial light, we increasingly have to move farther away from towns and cities to see a sky full of stars.
Light pollution poses a challenge for nocturnal species that have adapted to living in the dimness of night (1, 2) — see documentaries about the impacts of artificial light on wildlife and insects, and a related scientific talk. This problem might be one of the causes of the global decline in insects (3, 4), in turn negatively affecting their role in maintaining agricultural systems through pest control, pollination, and soil quality (5). These concepts are featured by the documentaries The Insect Apocalypse and The Great Death of Insects.
Chocó golden scarab (Chrysina argenteola) walking on forest litter in La Maná (Cotopaxi, Ecuador). Growing to up to 4 cm in length, this species inhabits the tropical rainforest of the Chocó region in the Western Andes (10), where it is frequently attracted to artificial lights at night. The striking colour of this ‘jewel scarab’ is an optical illusion. The exoskeleton is covered with overlapping layers of chitin that polarise light and reflect hues of blue, gold, green, silver, or reddish tones, depending on the species (16). The metallic sheen appears to deter bird predation (17) and might serve as camouflage as well as aid in individual recognition (11). The eyes of insects are ‘compound’ — composed of 100s to 1000s of tubular eyelets (‘ommatidia’), each with its own cornea and lens (18), and all collectively contributing to insect vision. In nocturnal species like the golden scarab, the photoreceptor cells (at the base of each ommatidium) respond more slowly to light compared to diurnal species, allowing the former to collect more nocturnal light per unit of time before forming an image (19). However, just as staring at the sun blinds us, eyes adapted for night vision become overwhelmed by excessive artificial light, disrupting the behaviour of these species. Below the scarab image are two photographs contrasting the day and night landscapes of the same location in Pedro Vicente Maldonado (Pichincha, Ecuador) within the species’ distribution range. Photos courtesy of Martín Bustamante (animal) and Luis Camacho (city).
When flying, nocturnal insects orient their backs toward the sky, using the light of the moon and stars as a reference (6) (explained here and here). However, when they encounter artificial lights, they can no longer distinguish up from down, and so they can become disoriented, flying erratically, like a moth circling a streetlight.
It is estimated that a third of the insects attracted to artificial light die from collisions, burn injuries, exhaustion, and/or predation (7). In the tropics, finding countless dead insects at the base of urban lights is a common scene. Equally important is that artificial light also hinders migration, foraging, and the search for mates in many nocturnal species (1, 8, 9).
Nocturnal jewels
Camacho and collaborators evaluated the effect of artificial lighting at night on the Chocó golden scarab (Chrysina argenteola) (10). This species inhabits the tropical rainforests of the Western Andes from Ecuador to Colombia, and is a member of the group known as ‘jewel scarabs‘ due to their metallic body coloration (11). Because of its nocturnal habits and the larvae’s dependence on wood for food (12), the golden scarab has been increasingly affected by the loss of native forest in combination with light pollution from rural and urban expansion.
In Australia, most fire occurs in the vast tropical savannas of the country’s north. In new research published in Nature Geoscience, we show Indigenous management of fire in these regions began at least 11,000 years ago – and possibly as long as 40,000 years ago.
But climate change and other effects of human activity are making wildfires more common and more severe in many regions, often with catastrophic results. In Australia, fires have caused major economic, environmental and personal losses, most recently in the south of the country.
Following my annual tradition, I present the retrospective list of the ‘top’ 20 influential papers of 2022 as assessed by experts in Faculty Opinions(formerly known as F1000). These are in no particular order. See previous years’ lists here: 2021, 2020, 2019, 2018, 2017, 2016, 2015, 2014, and 2013.
The logic of money contradicts the logic of species conservation and human health. As illegal trade has driven pangolins to near extinction, their hunting and market value has kept increasing ― even when we have known that they act as coronavirus reservoirs in the middle of the Covid-19 pandemic.
Sunda pangolin (Manis javanica) in a monsoon forest (Sumba Island, Indonesia). With adult weights up to 10 kg and body lengths around half a metre, these animals are mostly solitary and nocturnal, feed on ants and termites, and love tree climbing using bark hollows to shelter and give birth to singletons. The species occurs across mainland and islands of South East Asia, and became ‘Endangered’ in 2008 and ‘Critically Endangered’ in 2014, following a 80% decline in the last 20 years due to hunting and poaching. It has been the most heavily trafficked Asian species, and the IUCN’s assessment states: “… the incentives for harvesting and illegally trading in the species are universally high based on the high financial value of pangolin parts and derivatives”. Captive breeding is unlikely to deter wild collection because (among other reasons) farming costs are high (more so on a large scale) and, even if the species could be traded legally, wild versus farmed pangolin products and individuals are difficult to distinguish (23). Photo courtesy of Michael Pitts
Urbanites are attracted to exotic species, materials, and places. Our purchasing power seems to give us the right to buy any ‘object’ that we can pay for, no matter how exotic the object might be. In such a capitalist rationale, it is no surprise that > 150 thousand illegal cargos with wild animals and plants have been confiscated in 149 countries over the last two decades, moving some 6000 species from one place of the planet to another (1).
Social networks show people interacting with all kinds of fauna, creating the illusion that any animal can become a pet (2). And there’s a multi-$billion market of wildlife for a diverse array of uses including collecting, food, ornamentation, leisure, clothing and medicine (3-5). The paradox is that the rarer a species is, the higher its market value runs and the more lucrative selling it turns out to be, leading to more exploitation and rocketing extinction risk (6).
The intensity of threats to biodiversity from human endeavour becomes weaker as the distance to them increases.
As you move away from the big city to enjoy the countryside, you’ll notice the obvious increase in biodiversity. Even the data strongly support this otherwise subjective perception — there is a positive correlation between the degree we destroy habitat, harvest species, and pollute the environment, and the distance from big cities.
Remote locations are therefore usually considered safe havens and potential reservoirs for biodiversity. But our new study published recently in Nature Communications shows how this obvious pattern depicts only half of the story, and that global conservation management and actions might benefit from learning more about the missing part.
Communities are not just lists of individual species. Instead, they consist of complex networks of ecological interactions linking interdependent species. The structure of such networks is a fundamental determinant of biodiversity emergence and maintenance. However, it also plays an essential role in the processes of biodiversity loss. The decline or disappearance of some species might have detrimental —often fatal — effects on their associates. For example, a parasite cannot survive without its hosts, as much as a predator will starve without prey, or a plant will not reproduce without pollinators.
Events where a species disappears following the loss of other species on which it depends are known as co-extinctions, and they are now recognised as a primary driver of the ongoing global biodiversity crisis. The potential risk stemming from ecological dependencies is a major concern for all ecological systems.
In our latest study, we examine the downstream effects of publicising an elevated species description for a reptile that is highly prized in the international commercial wildlife trade.
We describe how iguanas from an insular population of the common green iguana (Iguana iguana) entered commercial trade shortly after an announcement was made indicating that the population would be described as a new species.
The international commercial wildlife trade presents a known risk factor for wild populations of threatened species. One organisation in particular regulates the international trade in species — the Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES).
Although most people probably know about the illegal practices involving iconic elephants and rhinos, reptiles are also targeted and traded. For example, after its discovery and description in 2016, and even though locality data were safeguarded, China’s endemic Mountain spiny crocodile newt (Echinotriton maxiquadratus) quickly entered the trade. This put conservation pressure on this small-range species (1, 2). Therefore, CITES signatory countries placed this species on its Appendix II in 2019, which lists animals and plants in need of protection.
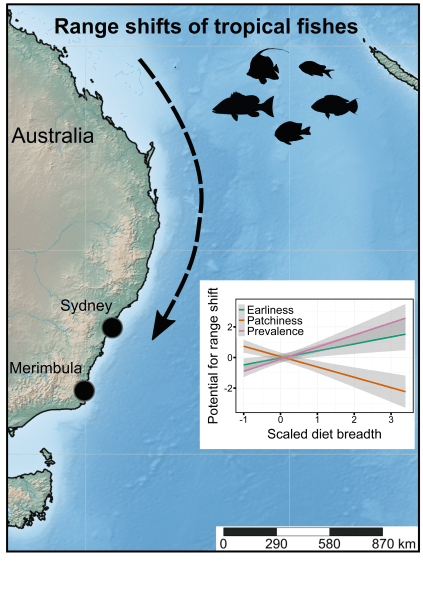
Ecologists often rely on measuring certain elements of a species’ characteristics, behaviour, or morphology to determine if these — what we call ‘traits’ — give them certain capacities to exploit their natural environments. While sometimes a bit arbitrarily defined, the traits that can be measured are many indeed, and sometimes they reveal rather interesting elements of a species’ resilience in the face of environmental change.
As we know, climate change is changing the way species are distributed around the planet, for the main (and highly simplified) reason that the environments in which they’ve evolved and to which they have adapted are changing.
In the simplest case, a warming climate means that there is a higher and higher chance you’ll experience temperatures that really don’t suit you that well (think of a koala or a flying fox baking in a tree when the thermometer reads +45° in the shade). Just like you seeking those nice, air-conditioned spaces on a scorcher of a day, species like to move to where conditions are more acceptable to their particular physiologies and behaviours.
Ecologists use life-history traits to predict which species have the highest probability of moving to new areas in response to climate change. Most studies into this phenomenon have largely ignored that range shifts in fact occur in sequential stages: (1) the species arrives in a new place for the first time, (2) its population increases in size (and extent), and (3) it can continue to persist in the new spot. Read the rest of this entry »
Rain forest gives way to pastures in the Brazilian Amazon in Mato Grosso. Photo by Thiago Foresti.
More than 600 scientists from every country in the EU and 300 Brazilian Indigenous groups have come together for the first time. This is because we see a window of opportunity in the ongoing trade negotiations between the EU and Brazil. In a Letter published in Science today, we are asking the EU to stand up for Brazilian Indigenous rights and the natural world. Strong action from the EU is particularly important given Brazil’s recent attempts to dismantle environmental legislation and ‘develop the unproductive Amazon’.
It’s worth clarifying — this isn’t about the EU trying to control Brazil — it’s about making sure our imports aren’t driving violence and deforestation. Foreign white people trying to ‘protect nature’ abroad have a dark and shameful past, where actions done in the name of conservation have led to the eviction of millions of Indigenous people. This has predominantly been to create (what we in the world of conservation would call) ‘protected areas’. The harsh reality is that most protected areas either are or have been ancestral lands of Indigenous people who are closely linked to their land and depend on it for their survival. Clearly, conservationists need to support Indigenous people. This new partnership between European scientists and Brazilian Indigenous groups is doing just that.
Brazil’s forest loss 2001-2013 shown in red. Indigenous lands outlined. By Mike Clark; data from GlobalForestWatch.org
In Brazil, many Indigenous groups still have a right to their land. This land is predominantly found in the Amazon rainforest, where close to a million Indigenous people live and depend on a healthy forest. Indigenous people are some of the best protectors of this vast forest, and are crucial to a future of long-term successful conservation. But Brazilian Indigenous groups and local communities are increasingly under attack. Violence on deforestation frontiers in Brazil has spiked this month, with at least 9 people found dead. The future is particularly scary for Indigenous people when there are quotes such as this from the man who is currently the President “It’s a shame that the Brazilian cavalry hasn’t been as efficient as the Americans, who exterminated the Indians.”
On top of human rights and environmental concerns, there is a strong profit driven case for halting deforestation. For example, ongoing deforestation in the Amazon risks flipping large parts of the rainforest to savanna – posing a serious risk to agricultural productivity, food security, local livelihoods, and the Brazilian economy. Zero-deforestation doesn’t harm agri-business, it allows for its longevity.Read the rest of this entry »
This week sees the launch of an updated bat synopsis from Conservation Evidence, adding new studies that have come out since the first synopsis was published in 2013.
The synopsis collects and summarises studies that test conservation actions such as ‘provide bat boxes for roosting bats’, and organises the studies by the action that they test. This focus on solutions makes it a handy point of reference for conservationists wishing to see what might work — and what is unlikely to work — to conserve bats.
Bechstein’s bat (Myotis bechsteinii) — photo credit Claire Wordley
Free to read or download from Conservation Evidence, the update represents a major addition to the original, containing 173 studies to the original 101. Studies are included if they tested an action that could be put in place for conservation, and measured an outcome for bats. As well as adding studies published from 2013 on, the update adds studies originally published in Spanish or Portuguese, and it is hoped that more languages will be added in future editions. Read the rest of this entry »
What do you say to a man whose list of conservation awards reads like a Star Wars film intro, who has introduced terms like the ‘hyperdynamism hypothesis’ to the field of ecology, and whose organisation reaches over one million people each week with updates of the scientific kind?
—
Well, I started by asking what it is that leads him to love the natural world to the extent he does. His answer was disarmingly simple.
“I grew up in the country, on an Oregon cattle ranch, and I think my love of nature just evolved naturally from that. When I was a young kid my dad and I did some fishing and ‘rock-hounding’— searching for rare stones and fossils. As an adolescent and teen I loved heading off into a forest or wilderness, rifle in hand – back in those days you could do that – to see whatever I could find. I watched red foxes hunting, eagles mating, and even heard a mountain lion scream. I got to be a pretty good duck and game-bird hunter.”
He’s quick to point out, however, he realised his taste for guns was not so developed as his love of nature.
“I gave up my rifles for a camera, and enjoyed that even more. I really got into photography for a while. Nature has always just calmed and fascinated me —I guess that’s partly why I became a conservationist.”
Who is Bill Laurance?
William F. Laurance is one of the leading ecology and conservation scientists globally, publishing dozens of papers in journals like Nature and Science, and rewriting the way scientists in the field research the complex interactions between flora and fauna — particularly in rainforests like the Amazon.
He is a Distinguished Research Professor at James Cook University in Australia, a Fellow of the Australian Academy of Science and the American Association for the Advancement of Science, and has received an Australian Laureate Fellowship from the Australian Research Council.
All this for a man from western USA who dreamed of running a zoo. Instead, he has travelled a path of intricate and game-changing research, trailblazing awareness campaigns and inspirational writings that have driven the way many see the environment over the past few decades.
Despite this profile, Laurance gave some time to tell Conjour about his life, his passion and his aims. I asked him what — considering his impressive CV — the future holds.
I’ve just read an interesting paper published in late 2016 in Conservation Biology that had so far escaped my attention. But given my interest in African conservation recently (and some interesting research results on the determinants of environmental performance for that region should be coming soon out of our lab), the work caught my eye.
The paper by Bowker and colleagues asked a question that has been asked previously regarding the ‘effectiveness’ of protected areas — do they succeed in limiting forest loss? While forest loss itself is not necessarily indicative of biodiversity erosion in any given area (for that, you need measures of species trends, etc.), it is arguably one of the most important drivers of species loss today.
The first set out to differentiate ‘effective’ from ‘ineffective’ protected areas, which was a simple binary variable related to whether there was less deforestation inside the protected area relative to comparable points outside (effective), or greater than or equal to deforestation outside (ineffective). The authors then related this binary response to a series of biophysical and social indicators. Read the rest of this entry »
Concrete jungle. A Barbary macaque sits in a human-dominated landscape in Gibraltar. Photo: Silviu Petrovan
Saving primates is a complicated business. Primates are intelligent, social animals that have complex needs. They come into conflict with humans when they raid rubbish bins and crops, chew power cables, and in some cases become aggressive towards people.
Humans, however, have the upper hand. While 60% of non-human primate species are threatened, humans grow in numbers and power, building roads through forests, hunting and trapping primates, and replacing their habitat with farms and houses.
To help primatologists choose the most effective conservation approaches to resolve these problems, researchers in the Conservation Evidence project teamed up with primate researchers to produce a global database on the effectiveness of primate conservation solutions. This free database, which can also be downloaded as a single pdf, summarizes the evidence for 162 conservation interventions — actions that conservationists might take to conserve primates. The data come from searches of over 170 conservation journals and newsletters, and each study is summarized in a single paragraph in plain English, making it possible for conservationists without access to scientific journals to read the key findings.
So what works in primate conservation? Well, the picture is rarely straightforward — partly due to the lack of data — but there are some interesting trends. Reducing hunting is one area where there seem to be a range of potentially effective approaches. Community control of patrolling, banning hunting and removing snares was effective in the three studies in which it was tested, all in African countries.
Thermal microhabitats are often uncoupled from above-ground air temperatures. A study focused on small frogs and lizards from the Philippines demonstrates that the structural complexity of tropical forests hosts a diversity of microhabitats that can reduce the exposure of many cold-blooded animals to anthropogenic climate warming.
Reproductive pair of the Luzon forest frogs Platymantis luzonensis (upper left), a IUCN near-threatened species restricted to < 5000 km2 of habitat. Lower left: the yellow-stripped slender tree lizard Lipinia pulchella, a IUCN least-concerned species. Both species have body lengths < 6 cm, and are native to the tropical forests of the Philippines. Right panels, top to bottom: four microhabitats monitored by Scheffers et al. (2), namely ground vegetation, bird’s nest ferns, phytotelmata, and fallen leaves above ground level. Photos courtesy of Becca Brunner (Platymantis), Gernot Kunz (Lipinia), Stephen Zozaya (ground vegetation) and Brett Scheffers (remaining habitats).
If you have ever entered a cave or an old church, you will be familiar with its coolness even in the dog days of summer. At much finer scales, from centimetres to millimetres, this ‘cooling effect’ occurs in complex ecosystems such as those embodied by tropical forests. The fact is that the life cycle of many plant and animal species depends on the network of microhabitats (e.g., small crevices, burrows, holes) interwoven by vegetation structures, such as the leaves and roots of an orchid epiphyte hanging from a tree branch or the umbrella of leaves and branches of a thick bush.
Much modern biogeographical research addressing the effects of climate change on biodiversity is based on macroclimatic data of temperature and precipitation. Such approaches mostly ignore that microhabitats can warm up or cool down in a fashion different from that of local or regional climates, and so determine how species, particularly ectotherms, thermoregulate (1). To illustrate this phenomenon, Brett Schefferset al. (2) measured the upper thermal limits (typically known as ‘critical thermal maxima’ or CTmax) of 15 species of frogs and lizards native to the tropical forest of Mount Banahaw, an active volcano on Luzon (The Philippines). The > 7000 islands of this archipelago harbour > 300 species of amphibians and reptiles (see video here), with > 100 occurring in Luzon (3).
I’ve recently read perhaps the most comprehensive treatise of forest fragmentation research ever compiled, and I personally view this rather readable and succinct review by Bill Laurance and colleagues as something every ecology and conservation student should read.
The ‘Biological Dynamics of Forest Fragments Project‘ (BDFFP) is unquestionably one of the most important landscape-scale experiments ever conceived and implemented, now having run 38 years since its inception in 1979. Indeed, it was way ahead of its time.
Experimental studies in ecology are comparatively rare, namely because it is difficult, expensive, and challenging in the extreme to manipulate entire ecosystems to test specific hypotheses relating to the response of biodiversity to environmental change. Thus, we ecologists tend to rely more on mensurative designs that use existing variation in the landscape (or over time) to infer mechanisms of community change. Of course, such experiments have to be large to be meaningful, which is one reason why the 1000 km2 BDFFP has been so successful as the gold standard for determining the effects of forest fragmentation on biodiversity.
Three of the coral species studied by Muir (2): (a) Acropora pichoni: Pohnpei Island, Pacific Ocean — deep-water species/IUCN ‘Near threatened’; (b) Acropora divaricate: Maldives, Indian ocean — mid-water species/IUCN ‘Near threatened’; and (c) Acropora gemmifera: Orpheus Island, Australia — shallow-water species/IUCN ‘Least Concern’. The IUCN states that the 3 species are vulnerable to climate change (acidification, temperature extremes) and demographic booms of the invading predator, the crown-of-thorns starfishAcanthaster planci. Photos courtesy of Paul Muir.
Global warming of the atmosphere and the oceans is modifying the distribution of many plants and animals. However, marine species are bound to face non-thermal barriers that might preclude their dispersal over wide stretches of the sea. Sunlight is one of those invisible obstacles for corals from the Indian and Pacific Oceans.
If we were offered a sumptuous job overseas, our professional success in an unknown place could be limited by factors like cultural or linguistic differences that have nothing to do with our work experience or expertise. If we translate this situation into biodiversity terms, one of the best-documented effects of global warming is the gradual dispersal of species tracking their native temperatures from the tropics to the poles (1). However, as dispersal progresses, many species encounter environmental barriers that are not physical (e.g., a high mountain or a wide river), and whose magnitude could be unrelated to ambient temperatures. Such invisible obstacles can prevent the establishment of pioneer populations away from the source.
Corals are ideal organisms to study this phenomenon because their life cycle is tightly geared to multiple environmental drivers (see ReefBase: Global Information System for Coral Reefs). Indeed, the growth of a coral’s exoskeleton relies on symbiotic zooxanthellae (see video and presentation), a kind of microscopic algae (Dinoflagellata) whose photosynthetic activity is regulated by sea temperature, photoperiod and dissolved calcium in the form of aragonite, among other factors.
Clown fish couple (Amphiprion percula) among the tentacles of anemone Heteractis magnifica in Kimbe Bay (Papua New Guinea) – courtesy of Mark McCormick. Clownfish protect anemones from predators and parasites in exchange of shelter and food. The fish tolerates the host’s venom because its skin is protected by a mucus layer some 2-3× thicker than phylogenetically related species (12); clownfish fabricate the mucus themselves and seem to obtain anemone antigens through a period of acclimation (13), but whether protection is acquired or innate is still debated. Clownfish are highly social bony fish, forming groups with one reproductive pair (up to 11 cm in length each) and several smaller, non-reproductive males. Reproduction is protandrous (also known as sequential hermaphroditism), so larvae are born male and, as soon as the reproductive female dies, her widower becomes female and the largest of the subsidiary males becomes the alpha male. The IUCN lists clownfish, generically named ‘anemone fish’, as threatened by the pet-trade industry and habitat degradation, although surprisingly, only 1 species has been assessed (A. sandaracinos). The clown anemone fish A. ocellaris is the species that inspired Nemo in the 2003 Academy-Award fiction movie – contrary to the logical expectation that the Oscars Red Carpet would generate support for conservation on behalf of Hollywood, of the 1568 species represented in the movie, only 16 % of those evaluated are threatened (14).
Smell is like noise, the more scents we breathe in one sniff, the more difficult it is to distinguish them to the point of olfactory saturation. Experimental work with clownfish reveals that the increase in dissolved carbon dioxide in seawater, mimicking ocean acidification, alters olfactory physiology, with potential cascading effects on the demography of species.
Places such as a restaurant, a hospital or a library have a characteristic bouquet, and we can guess the emotional state of other people by their scents. Smell is critical between predators and prey of many species because both have evolved to detect each other without the aid of vision. At sea, the smell of predators dissolves in water during detection, attack, capture, and ingestion of prey, and many fishes use this information to assess the risk of ending up crunched by enemy teeth (1, 2). But predator-prey interactions can be modified by changes in the chemical composition of seawater and are therefore highly sensitive to ongoing ocean acidification (see global measuring network here). Experts regard ocean acidification as the ‘other CO2 problem’ of climate change (3) — just to emphasize that anthropogenic climate-change impacts terrestrial and aquatic ecosystems alike. Acidification occurs because the ocean absorbs CO2 at a rate proportional with the concentration of this gas in the atmosphere and, once dissolved, CO2 becomes carbonic acid (H2CO3), which in turn releases protons (H+) — in simple terms, pH is the concentration of protons (see video about ocean acidification): Read the rest of this entry »
Coral reef fishes are wonderfully diverse in size, form, and function, as well as their need for different habitats throughout the life cycle. Some species spend all of their life in the same kind of coral habitat, while others need different places to breed and feed.
Fishes requiring different habitats as they progress through life often have what we call ‘nurseries’ in which adults lay eggs and the subsequent juveniles remain, and these places are often dominated by mangroves or seagrasses (i.e., they are not part of the coral reef).
While we’ve known for quite some time that when these nursery habitats are not around, adjacent coral reefs have few, if any, of these nursery-dependent species. What we haven’t known until now is just how far the influence of nurseries extends along a coral reef.
In other words, if a nursery is present, just how many new recruits do different areas of a reef receive from it? Read the rest of this entry »
Common coqui frog male (Eleutherodactylus coqui, snout-to vent length average ~ 3 cm) camouflaged in the fronds of an epiphyte in the El Yunque National Forest (Puerto Rico), along with an image of the enchanted forest of the Sierra de Luquillo where Narins & Meenderink did their study (4) – photos courtesy of Thomas Fletcher. This species can be found from sea level to the top of the highest peak in Puerto Rico (Cerro Punta = 1338 m). Native to mesic ecosystems, common coquis are well adapted to a terrestrial life, e.g., they lack interdigital webbing that support swimming propulsion in many amphibians, and youngsters hatch directly from the egg without transiting a tadpole stage. The IUCN catalogues the species as ‘Least Concern’ though alerts recent declines in high-altitude populations caused by chytrid fungus – lethal to amphibians at a planetary scale (9). Remarkably, the species has been introduced to Florida, Hawaii, the Dominican Republic and the Virgin Islands where it can become a pest due to high fertility rates (several >20 egg clutches/female/year).
Frog songs are species-specific and highly useful for the study of tropical communities, which host the highest amphibian diversities globally. The auditory system of females and the vocal system of males have co-evolved to facilitate reproductive encounters, but global warming might be disrupting the frequency of sound-based encounters in some species..
Amphibians (see Amphibians for kids by National Geographic) also love to sing in rainy nights when males call for a partner, but now they have to do it in hotter conditions as local climates become warmer. Vocal behaviour is a critical trait in the life history of many frog species because it mediates recognition between individuals, including sexual selection by females (1).
With few exceptions, every species has a different and unique call, so scientists can use call features to identify species, and this trait is particularly useful in the inventory of diverse tropical communities (2). Differences in call frequency, duration and pitch, and in note, number, and repetition pattern, occur from one species to another. And even within species, songs can vary from individual to individual (as much as there are not two people with the same voice), and be tuned according to body size and environmental temperature (3). Read the rest of this entry »
I’ve recently come across an interesting study that perfectly marries palaeo-ecological data with modern conservation philosophy. It’s not often that such a prehistorical perspective dating at least to the Last Glacial Maximum has been used so effectively to inform future conservation outlooks. I’m particularly interested in this sort of approach considering my own palaeo dabblings of late.
Published in Nature Communications this May, Lydia Cole and colleagues’ paper Recovery and resilience of tropical forests after disturbance is a meta-analysis of 71 studies covering nearly 300 disturbance events in tropical forests over the last 20,000 years or so. Using fossil pollen records as an index of vegetation change, they demonstrated the (somewhat intuitive) main result that the time to recovery following a disturbance generally decreases as the past disturbance frequency increased.
This appears to be a vindication of the idea that a system’s adaptive strategies evolve as a product of the local disturbance regime. More importantly, they found that recovery was faster following ‘large infrequent events’, which are natural perturbations such as cyclones and major fires. While most past disturbances were caused by humans clearing forest, the fact that tropical forest systems were most resilient to ‘natural’ events means that if we can’t stop human disturbances, at least we can attempt to emulate natural processes to maximise the rebound potential. Much like many modern forestry operations try to emulate natural disturbances to limit their damage, we should at least manage our impacts by understanding so-called ‘natural’ regimes as much as possible. Read the rest of this entry »
Just over two years ago I reported the ‘likely’ eradication of feral pigs from Australia’s third-largest (4,405 km2) island — Kangaroo Island. I indicated ‘likely’ because the program still required the proof-of-eradication phase to be completed before an official declaration could be made. Yesterday I had the immense honour to take part in the official…
Have you ever done any research that relied to any degree on Indigenous Knowledges? How did you cite those Knowledges, if at all? It’s probably time we rethink how we engage with Indigenous Knowledge systems. In a new article published in BioScience, we — a large group of Indigenous and non-Indigenous scholars in Australia —…
A recent paper, co-authored with the late Paul Ehrlich, reveals that the global human population has surpassed Earth’s sustainable capacity. It highlights the dire implications for food security, climate stability, and wellbeing. The study underscores that immediate changes in consumption and population management are crucial for a sustainable future.








 Ecologists often rely on measuring certain elements of a species’ characteristics, behaviour, or morphology to determine if these — what we call ‘traits’ — give them certain capacities to exploit their natural environments. While sometimes a bit arbitrarily defined, the traits that can be measured are many indeed, and sometimes they reveal rather interesting elements of a species’ resilience in the face of environmental change.
Ecologists often rely on measuring certain elements of a species’ characteristics, behaviour, or morphology to determine if these — what we call ‘traits’ — give them certain capacities to exploit their natural environments. While sometimes a bit arbitrarily defined, the traits that can be measured are many indeed, and sometimes they reveal rather interesting elements of a species’ resilience in the face of environmental change.

 What do you say to a man whose list of conservation awards reads like a Star Wars film intro, who has introduced terms like the ‘hyperdynamism hypothesis’ to the field of ecology, and whose organisation reaches over one million people each week with updates of the scientific kind?
What do you say to a man whose list of conservation awards reads like a Star Wars film intro, who has introduced terms like the ‘hyperdynamism hypothesis’ to the field of ecology, and whose organisation reaches over one million people each week with updates of the scientific kind?














{kind=link}