Using animals as sport symbols reflects the integration of biodiversity into cultural identity and the transmission of collective values. This raises the possibility that the economic muscle of the sport industry could translate its symbolic capital into tangible commitments to biodiversity conservation.
Those who have had the privilege of travelling in remote areas might have come across an unexpected scene: a football pitch in the middle of the Amazon rain forest or on the slopes of the Andes, a basketball court on the side of a Buddhist temple, or an ice hockey rink on a snow-ploughed lake in remote northern Canada.
Sport is a global industry that generates identity, belonging, education, and shared emotions for both athletes and their avid spectators (1). Sporting affinities now rival the sense of nationhood once shared by citizens during warfare (2).
Now in our heavily monetised world, sport clubs rely on their fans through tickets and merchandising, and indirectly through television rights and advertising. In this both emotional and commercial relationship, expressions such as being true to the badge reflect the central role of corporate symbols in building bonds between a club and its supporters (3).

In professional sport such as football, clubs increasingly function as brands (4) where even traditional logos are modified to enhance a team’s commercial value and strengthen audience loyalty (5). In this process, biodiversity becomes relevant because the iconography of many sport organisations incorporates representations of plants and animals.
Sport fauna
To quantify this phenomenon, Ugo Arbieu and collaborators analysed the presence of animals in club names, crests, and fan nicknames among 10 professional team sports across 50 countries (6). They found that 727 teams use 161 different animal species in their corporate imagery. Football and basketball lead in the number of species represented due to the large number of clubs worldwide, but American football, rugby and baseball display greater symbolic fauna diversity per club. Mammals and birds are the most common, particularly carnivores and raptors.
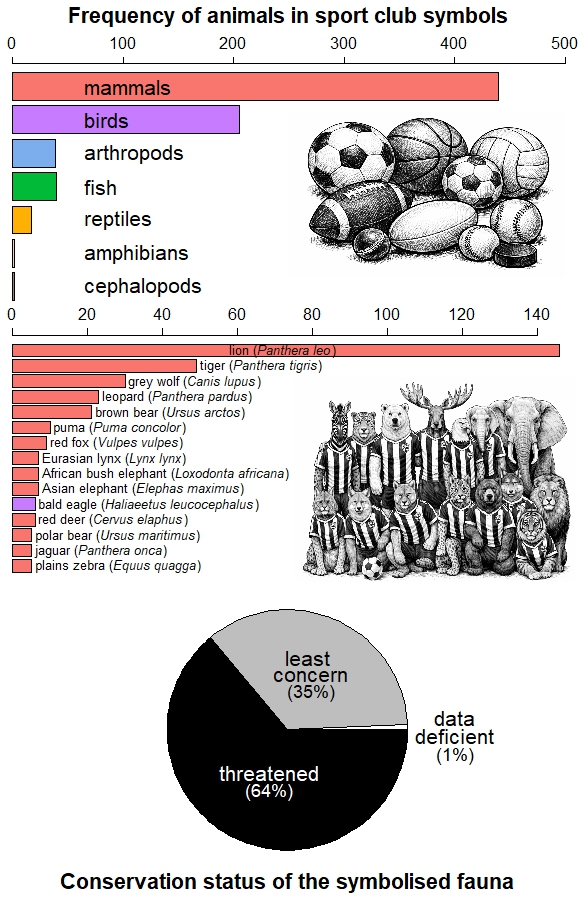
This pattern is not coincidental, for it reflects the historical bias of science and conservation towards large, charismatic vertebrates (7), but also the uneven availability of biological information and our social preferences for certain species (8). These preferences are even reflected in the animal emojis we share regularly on social media (9).
Arbieu’s study also revealed that clubs tend to favour images of threatened fauna (6), possibly due to their higher symbolic impact and media visibility (10). Moreover, although clubs in Europe and the Americas more often depict exotic animals, native species dominate in Africa, Asia, and Oceania (6). This suggests that the choice of an animal as an emblem is the product of not only aesthetic or symbolic criteria, but also of cultural roots and the historical relationship of societies with their local fauna.
Read the rest of this entry »










 it’s a provocative title, I agree. But then again, it’s true.
it’s a provocative title, I agree. But then again, it’s true.






 The global human population is now over 7.5 billion, and increasing by about
The global human population is now over 7.5 billion, and increasing by about 




