Flooding in the Murray-Darling Basin is creating ideal breeding conditions for many native species that have evolved to take advantage of temporary flood conditions. Led by PhD candidate Rupert Mathwin, our team developed virtual models of the Murray River to reveal a crucial link between natural flooding and the extinction risk of endangered southern bell frogs (Litoria raniformis; also known as growling grass frogs).
Southern bell frogs are one of Australia’s 100 Priority Threatened Species. This endangered frog breeds during spring and summer when water levels increase in their wetlands. However, the natural flooding patterns in Australia’s largest river system have been negatively impacted by expansive river regulation that some years, sees up to 60% of river water extracted for human use.
Our latest paper describes how we built computer simulations of Murray-Darling Basin wetlands filled with simulated southern bell frogs. By changing the simulation from natural to regulated conditions, we showed that modern conditions dramatically increase the extinction risk of these beloved frogs.
The data clearly indicate that successive dry years raise the probability of local extinction, and these effects are strongest in smaller wetlands. Larger wetlands and those with more frequent inundation are less prone to these effects, although they are not immune to them entirely. The models present a warning — we have greatly modified the way the river behaves, and the modern river cannot support the long-term survival of southern bell frogs.’
Last week, researchers at the University of Melbourne announced that thylacines or Tasmanian tigers, the Australian marsupial predators extinct since the 1930s, could one day be ushered back to life.
The thylacine (Thylacinus cynocephalus), also known as the ‘Tasmanian tiger’ (it was neither Tasmanian, because it was once common in mainland Australia, nor was it related to the tiger), went extinct in Tasmania in the 1930s from persecution by farmers and habitat loss. Art by Eleanor (Nellie) Pease, University of Queensland. Centre of Excellence for Australian Biodiversity and Heritage
Advances in mapping the genome of the thylacine and its living relative the numbat have made the prospect of re-animating the species seem real. As an ecologist, I would personally relish the opportunity to see a living specimen.
The announcement led to some overhyped headlines about the imminent resurrection of the species. But the idea of “de-extinction” faces a variety of technical, ethical and ecological challenges. Critics (like myself) argue it diverts attention and resources from the urgent and achievable task of preventing still-living species from becoming extinct.
The rebirth of the bucardo
The idea of de-extinction goes back at least to the the creation of the San Diego Frozen Zoo in the early 1970s. This project aimed to freeze blood, DNA, tissue, cells, eggs and sperm from exotic and endangered species in the hope of one day recreating them.
The notion gained broad public attention with the first of the Jurassic Park films in 1993. The famous cloning of Dolly the sheep reported in 1996 created a sense that the necessary know-how wasn’t too far off.
The next technological leap came in 2008, with the cloning of a dead mouse that had been frozen at –20℃ for 16 years. If frozen individuals could be cloned, re-animation of a whole species seemed possible.
After this achievement, de-extinction began to look like a potential way to tackle the modern global extinction crisis.
As someone who writes a lot of models — many for applied questions in conservation management (e.g., harvest quotas, eradication targets, minimum viable population sizes, etc.), and supervises people writing even more of them, I’ve had many different experiences with their uptake and implementation by management authorities.
Some of those experiences have involved catastrophic failures to influence any management or policy. One particularly painful memory relates to a model we wrote to assist with optimising approaches to eradicate (or at least, reduce the densities of) feral animals in Kakadu National Park. We even wrote the bloody thing in Visual Basic (horrible coding language) so people could run the module in Excel. As far as I’m aware, no one ever used it.
Others have been accepted more readily, such as a shark-harvest model, which (I think, but have no evidence to support) has been used to justify fishing quotas, and one we’ve done recently for the eradication of feral pigs on Kangaroo Island (as yet unpublished) has led directly to increased funding to the agency responsible for the programme.
According to Altmetrics (and the online tool I developed to get paper-level Altmetric information quickly), only 3 of the 16 of what I’d call my most ‘applied modelling’ papers have been cited in policy documents:
For many years I’ve been interested in modelling the extinction dynamics of megafauna. Apart from co-authoring a few demographically simplified (or largely demographically free) models about how megafauna species could have gone extinct, I have never really tried to capture the full nuances of long-extinct species within a fully structured demographic framework.
That is, until now.
But how do you get the life-history data of an extinct animal that was never directly measured. Surely, things like survival, reproductive output, longevity and even environmental carrying capacity are impossible to discern, and aren’t these necessary for a stage-structured demographic model?
The answer to the first part of that question “it’s possible”, and to the second, it’s “yes”. The most important bit of information we palaeo modellers need to construct something that’s ecologically plausible for an extinct species is an estimate of body mass. Thankfully, palaeontologists are very good at estimating the mass of the things they dig up (with the associated caveats, of course). From such estimates, we can reconstruct everything from equilibrium densities, maximum rate of population growth, age at first breeding, and longevity.
But it’s more complicated than that, of course. In Australia anyway, we’re largely dealing with marsupials (and some monotremes), and they have a rather different life-history mode than most placentals. We therefore have to ‘correct’ the life-history estimates derived from living placental species. Thankfully, evolutionary biologists and ecologists have ways to do that too.
The Pleistocene kangaroo Procoptodon goliah, the largest and most heavily built of the short-faced kangaroos, was the largest and most heavily built kangaroo known. It had an unusually short, flat face and forwardly directed eyes, with a single large toe on each foot (reduced from the more normal count of four). Each forelimb had two long, clawed fingers that would have been used to bring leafy branches within reach.
So with a battery of ecological, demographic, and evolutionary tools, we can now create reasonable stochastic-demographic models for long-gone species, like wombat-like creatures as big as cars, birds more than two metres tall, and lizards more than seven metres long that once roamed the Australian continent.
Ancient clues, in the shape of fossils and archaeological evidence of varying quality scattered across Australia, have formed the basis of several hypotheses about the fate of megafauna that vanished during a peak about 42,000 years ago from the ancient continent of Sahul, comprising mainland Australia, Tasmania, New Guinea and neighbouring islands.
There is a growing consensus that multiple factors were at play, including climate change, the impact of people on the environment, and access to freshwater sources.
Just published in the open-access journal eLife, our latest CABAH paper applies these approaches to assess how susceptible different species were to extinction – and what it means for the survival of species today.
Using various characteristics such as body size, weight, lifespan, survival rate, and fertility, we (Chris Johnson, John Llewelyn, Vera Weisbecker, Giovanni Strona, Frédérik Saltré & me) created population simulation models to predict the likelihood of these species surviving under different types of environmental disturbance.
We compared the results to what we know about the timing of extinction for different megafauna species derived from dated fossil records. We expected to confirm that the most extinction-prone species were the first species to go extinct – but that wasn’t necessarily the case.
While we did find that slower-growing species with lower fertility, like the rhino-sized wombat relative Diprotodon, were generally more susceptible to extinction than more-fecund species like the marsupial ‘tiger’ thylacine, the relative susceptibility rank across species did not match the timing of their extinctions recorded in the fossil record.
Indeed, we found no clear relationship between a species’ inherent vulnerability to extinction — such as being slower and heavier and/or slower to reproduce — and the timing of its extinction in the fossil record.
In fact, we found that most of the living species used for comparison — such as short-beaked echidnas, emus, brush turkeys, and common wombats — were more susceptible on average than their now-extinct counterparts.
However, this time I’ve strayed from my recent bibliometric musings and developed something that’s more compatible with the core of my main research and interests.
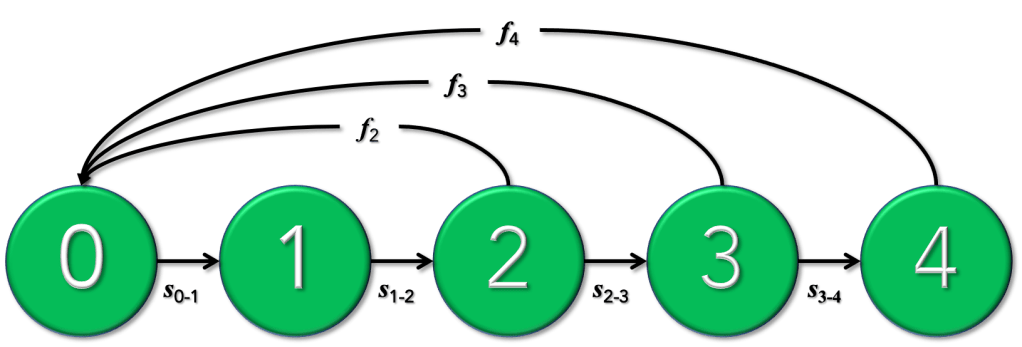
Over the years I’ve taught many students the basics of population modelling, with the cohort-based approaches dominating the curriculum. Of these, the simpler ‘Leslie’ (age-classified) matrix models are both the easiest to understand and for which data can often be obtained without too many dramas.
But unless you’re willing to sit down and learn the code, they can be daunting to the novice.
Sure, there are plenty of software alternatives out there, such as Bob Lacy‘s Vortex (a free individual-based model available for PCs only), Resit Akçakaya & co’s RAMAS Metapop ($; PC only), Stéphane Legendre‘s Unified Life Models (ULM; open-source; all platforms), and Charles Todd‘s Essential (open-source; PC only) to name a few. If you’re already an avid R user and already into population modelling, you might be familiar with the population-modelling packages popdemo, OptiPopd, or sPop. I’m sure there are still other good resources out there of which I’m not aware.
But, even to install the relevant software or invoke particular packages in R takes a bit of time and learning. It’s probably safe to assume that many people find the prospect daunting.
It’s for this reason that I turned my newly acquired R Shiny skills to matrix population models so that even complete coding novices can run their own stochastic population models.
One of the most ancient peopling events of the great diaspora of anatomically modern humans out of Africa more than 50,000 years ago — human arrival in the great continent of Sahul (New Guinea, mainland Australia & Tasmania joined during periods of low sea level) — remains mysterious. The entry routes taken, whether migration was directed or accidental, and just how many people were needed to ensure population viability are shrouded by the mists of time. This prompted us to build stochastic, age-structured human population-dynamics models incorporating hunter-gatherer demographic rates and palaeoecological reconstructions of environmental carrying capacity to predict the founding population necessary to survive the initial peopling of late-Pleistocene Sahul.
—
As ecological modellers, we are often asked by other scientists to attempt to render the highly complex mechanisms of entire ecosystems tractable for virtual manipulation and hypothesis testing through the inevitable simplification that is ‘a model’. When we work with scientists studying long-since-disappeared ecosystems, the challenges multiply.
Add some multidisciplinary data and concepts into the mix, and the complexity can quickly escalate.
This is how we tackled one of these big questions: just how did the first anatomically modern Homo sapiens make it to the continent and survive?
At that time, Australia was part of the giant continent of Sahul that connected New Guinea, mainland Australia, and Tasmania at times of lower sea level. In fact, throughout most of last ~ 126,000 years (late Pleistocene and much of the Holocene), Sahul was the dominant landmass in the region (see this handy online tool for how the coastline of Sahul changed over this period).
I published this last week on The Conversation, and now reproducing it here for CB.com readers.
—
Two days ago, the last male northern white rhino (Ceratotherium simum cottoni) died. His passing leaves two surviving members of his subspecies: both females who are unable to bear calves.
Even though it might not be quite the end of the northern white rhino because of the possibility of implanting frozen embryos in their southern cousins (C. simum simum), in practical terms, it nevertheless represents the end of a long decline for the subspecies. It also raises the question: how many individuals does a species need to persist?
Fiction writers have enthusiastically embraced this question, most often in the post-apocalypsegenre. It’s a notion with a long past; the Adam and Eve myth is of course based on a single breeding pair populating the entire world, as is the case described in the Ragnarok, the final battle of the gods in Norse mythology.
This idea dovetails neatly with the image of Noah’s animals marching “two by two” into the Ark. But the science of “minimum viable populations” tells us a different story.
With the official start of the new ARCCentre of Excellence for Australian Biodiversity and Heritage (CABAH) in July, I am pleased to announce two new CABAH-funded postdoctoral positions (a.k.a. Research Associates) in my global ecology lab at Flinders University in Adelaide (Flinders Modelling Node).
One of these positions is a little different, and represents something of an experiment. The Research Associate in Palaeo-Vegetation Modelling is being restricted to women candidates; in other words, we’re only accepting applications from women for this one. In a quest to improve the gender balance in my lab and in universities in general, this is a step in the right direction.
The project itself is not overly prescribed, but we would like something along the following lines of inquiry: Read the rest of this entry »
I have another postdoctoral fellowship to advertise! All the details you need for applying are below.
—
KEY PURPOSE
Scientific data such as fossil and archaeological records used as proxy to reconstruct past environments and biological communities (including humans) are sparse, often ambiguous or contradictory when establishing any consensus on timing or routes of initial human arrival and subsequent spread, the timing or extent of major changes in climate and other environmental perturbations, or the timing or regional pattern of biological extinctions.
The Research Fellow (Palaeo-Ecological Modelling) will assist in addressing these problems by developing state-of-the-art analytical and simulation tools to infer regional pattern of both the timing of human colonisation and megafauna extinction based on incomplete and sparse dataset, and investigating past environmental changes and human responses to identify their underlying causes and consequences on Australia’s landscapes, biodiversity and cultural history.
ORGANISATIONAL ENVIRONMENT
The position will be based in the School of Biological Sciences in the Faculty of Science & Engineering at Flinders University. Flinders University boasts a world-class Palaeontology Research Group (PRG) and the new Global Ecology Research Laboratory that have close association with the research-intensive South Australian Museum. These research groups contribute to building a dynamic research environment that explores the continuum of environmental and evolutionary research from the ancient to modern molecular ecology and phylogeography. The School of Biological Sciences is an integrated community researching and teaching biology, and has a long history of science innovation. The appointee will join an interdisciplinary school of approximately 45 academic staff. The teaching and research activities of the School are supported by a range of technical and administrative infrastructure services.
KEY RESPONSIBILITIES
The key responsibilities and selection criteria identified for this position should be read in conjunction with the Flinders University Academic Profiles for the relevant academic classification (scroll down to Academic Profiles).
The Research Fellow (Palaeo-Ecological Modelling) will work under the direction of the Project Chief Investigator, and will be required to: Read the rest of this entry »
In July 2015 an American dentist shot and killed a male lion called ‘Cecil’ with a hunting bow and arrow, an act that sparked a storm of social media outrage. Cecil was a favourite of tourists visiting Hwange National Park in Zimbabwe, and so the allegation that he was lured out of the Park to neighbouring farmland added considerable fuel to the flames of condemnation. Several other aspects of the hunt, such as baiting close to national park boundaries, were allegedly done illegally and against the spirit and ethical norms of a managed trophy hunt.
In May 2015, a Texan legally shot a critically endangered black rhino in Namibia, which also generated considerable online ire. The backlash ensued even though the male rhino was considered ‘surplus’ to Namibia’s black rhino populations, and the US$350,000 generated from the managed hunt was to be re-invested in conservation. Together, these two incidents have triggered vociferous appeals to ban trophy hunting throughout Africa.
These highly politicized events are but a small component of a large industry in Africa worth > US$215 million per year that ‘sells’ iconic animals to (mainly foreign) hunters as a means of generating otherwise scarce funds. While to most people this might seem like an abhorrent way to generate money, we argue in a new paper that sustainable-use activities, such as trophy hunting, can be an important tool in the conservationist’s toolbox. Conserving biodiversity can be expensive, so generating money is a central preoccupation of many environmental NGOs, conservation-minded individuals, government agencies and scientists. Making money for conservation in Africa is even more challenging, and so we argue that trophy hunting should and could fill some of that gap. Read the rest of this entry »
Did ice ages cause the Pleistocene megafauna to go extinct? Contrary to popular opinion, no, they didn’t. But climate change did have something to do with them, only it was global warming events instead.
Just out today in Science, our long-time-coming (9 years in total if you count the time from the original idea to today) paper ‘Abrupt warmings drove Late Pleistocene Holarctic megafaunal turnover‘ demonstrates for the first time that abrupt warming periods over the last 60,000 years were at least partially responsible for the collapse of the megafauna in Eurasia and North America.
You might recall that I’ve been a bit sceptical of claims that climate changes had much to do with megafauna extinctions during the Late Pleistocene and early Holocene, mainly because of the overwhelming evidence that humans had a big part to play in their demise (surprise, surprise). What I’ve rejected though isn’t so much that climate had nothing to do with the extinctions; rather, I took issue with claims that climate change was the dominant driver. I’ve also had problems with blanket claims that it was ‘always this’ or ‘always that’, when the complexity of biogeography and community dynamics means that it was most assuredly more complicated than most people think.
I’m happy to say that our latest paper indeed demonstrates the complexity of megafauna extinctions, and that it took a heap of fairly complex datasets and analyses to demonstrate. Not only were the data varied – the combination of scientists involved was just as eclectic, with ancient DNA specialists, palaeo-climatologists and ecological modellers (including yours truly) assembled to make sense of the complicated story that the data ultimately revealed. Read the rest of this entry »
Dick’s latest paper in Molecular Ecology is a meta-analysis designed to test whether there are any genetic grounds for NOT attempting genetic rescue for inbreeding-depressed populations. I suppose a few definitions are in order here. Genetic rescue is the process, either natural or facilitated, where inbred populations (i.e., in a conservation sense, those comprising too many individuals bonking their close relatives because the population in question is small) receive genes from another population such that their overall genetic diversity increases. In the context of conservation genetics, ‘inbreeding depression‘ simply means reduced biological fitness (fertility, survival, longevity, etc.) resulting from parents being too closely related.
Seems like an important thing to avoid, so why not attempt to facilitate gene flow among populations such that those with inbreeding depression can be ‘rescued’? In applied conservation, there are many reasons given for not attempting genetic rescue: Read the rest of this entry »
A major media release today coordinated by Jessica Meeuwig in Western Australia makes the (obvious) point that there’s no biological justification to cull sharks.
Coordinating scientist, Professor Jessica Meeuwig from the University of Western Australia said:
“To have over 300 researchers, including some of the world’s top shark specialists and marine ecologists, all strongly agreeing that there is no scientific basis for the lethal drum-line programme, tells you how unjustified the government’s proposal is. If the EPA and the Federal Minister for the Environment are using science for decisions, the drum-line proposal should not be approved.”
The experts agree that the proposal presents no evidence that the lethal drum-line programme, as implemented, will improve ocean safety. It ignores evidence from other hook-based programs in Hawaii and Queensland that have been shown to be ineffective in reducing shark attacks on humans.
“There is no evidence that drum lines reduce shark bites. The Western Australia EPA now faces a question of science versus politics with global implications because it is considering establishing a new international norm that would allow for the killing of protected white sharks.”
The drum lines are ineffective and indiscriminate, with 78% of the sharks captured not considered ‘threatening’ to humans. Yet, scientifically supported, non-lethal alternatives such as the South African ‘Shark Spotter’ and Brazil’s ‘Tag and Remove’ programmes are not adequately assessed as viable options for Western Australia. Read the rest of this entry »
As you might recall, Dick Frankham, Barry Brook and I recently wrote a review in Biological Conservationchallenging the status quo regarding the famous 50/500 ‘rule’ in conservation management (effective population size [Ne] = 50 to avoid inbreeding depression in the short-term, and Ne = 500 to retain the ability to evolve in perpetuity). Well, it inevitably led to some comments arising in the same journal, but we were only permitted by Biological Conservation to respond to one of them. In our opinion, the other comment was just as problematic, and only further muddied the waters, so it too required a response. In a first for me, we have therefore decided to publish our response on the arXivpre-print server as well as here on ConservationBytes.com.
—
50/500 or 100/1000 debate is not about the time frame – Reply to Rosenfeld
cite as: Frankham, R, Bradshaw CJA, Brook BW. 2014. 50/500 or 100/1000 debate is not about the time frame – Reply to Rosenfeld. arXiv: 1406.6424 [q-bio.PE]25 June 2014.
The Letter from Rosenfeld (2014) in response to Jamieson and Allendorf (2012) and Frankham et al. (2014) and related papers is misleading in places and requires clarification and correction, as follows: Read the rest of this entry »
A few weeks ago I wrote a post about how to run the perfect scientific workshop, which most of you thought was a good set of tips (bizarrely, one person was quite upset with the message; I saved him the embarrassment of looking stupid online and refrained from publishing his comment).
As I mentioned at the end of post, the stimulus for the topic was a particularly wonderful workshop 12 of us attended at beautiful Linnaeus Estate on the northern coast of New South Wales (see Point 5 in the ‘workshop tips’ post).
I hate to say it – mainly because it deserves as little attention as possible – but the main reason is that we needed to clean up a bit of rubbish. The rubbish in question being the latest bit of excrescence growing on that accumulating heap produced by a certain team of palaeontologists promulgating their ‘it’s all about the climate or nothing’ broken record.
Chiew Larn Reservoir is surrounded by Khlong Saeng Wildlife Sanctuary and Khao Sok National Park, which together make up part of the largest block of rainforest habitat in southern Thailand (> 3500 km2). Photo: Antony Lynam
One of the perennial and probably most controversial topics in conservation ecology is when is something “too small’. By ‘something’ I mean many things, including population abundance and patch size. We’ve certainly written about the former on many occasions (see here, here, here and here for our work on minimum viable population size), with the associated controversy it elicited.
Now I (sadly) report on the tragedy of the second issue – when is a habitat fragment too small to be of much good to biodiversity?
Published today in the journal Science, Luke Gibson (of No substitute for primary forest fame) and a group of us report disturbing results about the ecological meltdown that has occurred on islands created when the Chiew Larn Reservoir of southern Thailand was flooded nearly 30 years ago by a hydroelectric dam.
As is the case in many parts of the world (e.g., Three Gorges Dam, China), hydroelectric dams can cause major ecological problems merely by flooding vast areas. In the case of Charn Liew, co-author Tony Lynam of Wildlife Conservation Society passed along to me a bit of poignant and emotive history about the local struggle to prevent the disaster.
“As the waters behind the dam were rising in 1987, Seub Nakasathien, the Superintendent of the Khlong Saeng Wildlife Sanctuary, his staff and conservationist friends, mounted an operation to capture and release animals that were caught in the flood waters.
It turned out to be distressing experience for all involved as you can see from the clips here, with the rescuers having only nets and longtail boats, and many animals dying. Ultimately most of the larger mammals disappeared quickly from the islands, leaving just the smaller fauna.
Later Seub moved to Huai Kha Khaeng Wildlife Sanctuary and fought an unsuccessful battle with poachers and loggers, which ended in him taking his own life in despair in 1990. A sad story, and his friend, a famous folk singer called Aed Carabao, wrote a song about Seub, the music of which plays in the video. Read the rest of this entry »
This week has been all about biogeography for me. While I wouldn’t call myself a ‘biogeographer’, I certainly do apply a lot of the discipline’s techniques.
This week I’m attending the 2013 Association of Ecology’s (INTECOL) and British Ecological Society’s joint Congress of Ecology in London, and I have purposefully sought out more of the biogeographical talks than pretty much anything else because the speakers were engaging and the topics fascinating. As it happens, even my own presentation had a strong biogeographical flavour this year.
Although the species-area relationship (SAR) is only one small aspect of biogeography, I’ve been slightly amazed that after more than 50 years since MacArthur & Wilson’s famous book, our discipline is still obsessed with SAR.
I’ve blogged about SAR issues before – what makes it so engaging and controversial is that SAR is the principal tool to estimate overall extinction rates, even though it is perhaps one of the bluntest tools in the ecological toolbox. I suppose its popularity stems from its superficial simplicity – as the area of an (classically oceanic) island increases, so too does the total number of species it can hold. The controversies surrounding such as basic relationship centre on describing the rate of that species richness increase with area – in other words, just how nonlinear the SAR itself is.
Even a cursory understanding of maths reveals the importance of estimating this curve correctly. As the area of an ‘island’ (habitat fragment) decreases due to human disturbance, estimating how many species end up going extinct as a result depends entirely on the shape of the SAR. Get the SAR wrong, and you can over- or under-estimate the extinction rate. This was the crux of the palaver over Fangliang He (not attending INTECOL) & Stephen Hubbell’s (attending INTECOL) paper in Nature in 2011.
The first real engagement of SAR happened with John Harte’s maximum entropy talk in the process macroecology session on Tuesday. What was notable to me was his adamant claim that the power-law form of SAR should never be used, despite its commonness in the literature. I took this with a grain of salt because I know all about how messy area-richness data can be, and why one needs to consider alternate models (see an example here). But then yesterday I listened to one of the greats of biogeography – Robert Whittaker – who said pretty much the complete opposite of Harte’s contention. Whittaker showed results from one of his papers last year that the power law was in fact the most commonly supported SAR among many datasets (granted, there was substantial variability in overall model performance). My conclusion remains firm – make sure you use multiple models for each individual dataset and try to infer the SAR from model-averaging. Read the rest of this entry »
Given the popularity of certain prescriptive posts on ConservationBytes.com, I thought it prudent to compile a list of software that my lab and I have found particularly useful over the years. This list is not meant to be comprehensive, but it will give you a taste for what’s out there. I don’t list the plethora of conservation genetics software that is available (generally given my lack of experience with it), but if this is your chosen area, I’d suggest starting with Dick Frankham‘s excellent book, An Introduction to Conservation Genetics.
—
1. R: If you haven’t yet loaded the open-source R programming language on your machine, do it now. It is the single-most-useful bit of statistical and programming software available to anyone anywhere in the sciences. Don’t worry if you’re not a fully fledged programmer – there are now enough people using and developing sophisticated ‘libraries’ (packages of functions) that there’s pretty much an application for everything these days. We tend to use R to the exclusion of almost any other statistical software because it makes you learn the technique rather than just blindly pressing the ‘go’ button. You could also stop right here – with R, you can do pretty much everything else that the software listed below does; however, you have to be an exceedingly clever programmer and have a lot of spare time. R can also sometimes get bogged down with too much filled RAM, in which case other, compiled languages such as PYTHON and C# are useful.
2. VORTEX/OUTBREAK/META-MODEL MANAGER, etc.: This suite of individual-based projection software was designed by Bob Lacy & Phil Miller initially to determine the viability of small (usually captive) populations. The original VORTEX has grown into a multi-purpose, powerful and sophisticated population viability analysis package that now links to its cousin applications like OUTBREAK (the only off-the-shelf epidemiological software in existence) via the ‘command centre’ META-MODEL MANAGER (see an examples here and here from our lab). There are other add-ons that make almost any population projection and hindcasting application possible. And it’s all free! (warning: currently unavailable for Mac, although I’ve been pestering Bob to do a Mac version).
3. RAMAS: RAMAS is the go-to application for spatial population modelling. Developed by the extremely clever Resit Akçakaya, this is one of the only tools that incorporates spatial meta-population aspects with formal, cohort-based demographic models. It’s also very useful in a climate-change context when you have projections of changing habitat suitability as the base layer onto which meta-population dynamics can be modelled. It’s not free, but it’s worth purchasing. Read the rest of this entry »
—
On Friday, March 15 in Washington DC, National Geographic and TEDx are hosting a day-long conference on species-revival science and ethics. In other words, they will be debating whether we can, and should, attempt to bring extinct animals back to life – a concept some call “de-extinction”.
The debate has an interesting line-up of ecologists, geneticists, palaeontologists (including Australia’s own Mike Archer), developmental biologists, journalists, lawyers, ethicists and even artists. I have no doubt it will be very entertaining.
But let’s not mistake entertainment for reality. It disappoints me, a conservation scientist, that this tired fantasy still manages to generate serious interest. I have little doubt what the ecologists at the debate will conclude.
Once again, it’s important to discuss the principal flaws in such proposals.
Put aside for the moment the astounding inefficiency, the lack of success to date and the welfare issues of bringing something into existence only to suffer a short and likely painful life. The principal reason we should not even consider the technology from a conservation perspective is that it does not address the real problem – mainly, the reason for extinction in the first place.
Even if we could solve all the other problems, if there is no place to put these new individuals, the effort and money expended is a complete waste. Habitat loss is theprincipal driver of species extinction and endangerment. If we don’t stop and reverse this now, all other avenues are effectively closed. Cloning will not create new forests or coral reefs, for example. Read the rest of this entry »
Harvesting and habitat alteration reduce many populations to just a few individuals, and then often extinction. A widely recommended conservation action is to supplement those populations with new individuals translocated from other regions. However, crossing local and foreign genes can worsen the prospects of recovery.
We are all hybrids or combinations of other people, experiences and things. Let’s think of teams (e.g., engineers, athletes, mushroom collectors). In team work, isolation from other team members might limit the appearance of innovative ideas, but the arrival of new (conflictive) individuals might in fact destroy group dynamics altogether. Chromosomes work much like this – too little or too much genetic variability among parents can break down the fitness of their descendants. These pernicious effects are known as ‘inbreeding depression‘ when they result from reproduction among related individuals, and ‘outbreeding depression‘ when parents are too genetically distant.
Location of the two USA sites providing spawners of largemouth bass for the experiments by Goldberg et al. (3): the Kaskaskia River (Mississipi Basin, Illinois) and the Big Cedar Lake (Great Lakes Basin, Wisconsin). Next to the map is shown an array of three of the 72-litre aquaria in an indoor environment under constant ambient temperature (25 ◦C), humidity (60%), and photoperiod (alternate 12 hours of light and darkness). Photo courtesy of T. Goldberg.
Recent studies have revised outbreeding depression in a variety of plants, invertebrates and vertebrates (1, 2). An example is Tony Goldberg’s experiments on largemouth bass (Micropterus salmoides), a freshwater fish native to North America. Since the 1990s, the USA populations have been hit by disease from a Ranavirus. Goldberg et al. (3) sampled healthy individuals from two freshwater bodies: the Mississipi River and the Great Lakes, and created two genetic lineages by having both populations isolated and reproducing in experimental ponds. Then, they inoculated the Ranavirus in a group of parents from each freshwater basin (generation P), and in the first (G1) and second (G2) generations of hybrids crossed from both basins. After 3 weeks in experimental aquaria, the proportion of survivors declined to nearly 30% in G2, and exceeded 80% in G1 and P. Clearly, crossing of different genetic lineages increased the susceptibility of this species to a pathogen, and the impact was most deleterious in G2. This investigation indicates that translocation of foreign individuals into a self-reproducing population can not only import diseases, but also weaken its descendants’ resistance to future epidemics.
A mechanism causing outbreeding depression occurs when hybridisation alters a gene that is only functional in combination with other genes. Immune systems are often regulated by these complexes of co-adapted genes (‘supergenes’) and their disruption is a potential candidate for the outbreeding depression reported by Goldberg et al. (3). Along with accentuating susceptibility to disease, outbreeding depression in animals and plants can cause a variety of deleterious effects such as dwarfism, low fertility, or shortened life span. Dick Frankham (one of our collaborators) has quantified that the probability of outbreeding depression increases when mixing takes place between (i) different species, (ii) conspecifics adapted to different habitats, (iii) conspecifics with fixed chromosomal differences, and (iv) populations free of genetic flow with other populations for more than 500 years (2).
A striking example supporting (some of) those criteria is the pink salmon (Oncorhynchus gorbuscha) from Auke Creek near Juneau (Alaska). The adults migrate from the Pacific to their native river where they spawn two years after birth, with the particularity that there are two strict broodlines that spawn in either even or odd year – that is, the same species in the same river, but with a lack of genetic flow between populations. In vitro mixture of the two broodlines and later release of hybrids in the wild have shown that the second generation of hybrids had nearly 50% higher mortality rates (i.e., failure to return to spawn following release) when born from crossings of parents from different broodlines than when broodlines were not mixed (4).
Get Conservation Bytes by Email Enter your email address to receive new posts on conservation, biodiversity, and environmental science directly in your inbox. No need to visit the website regularly, and you can unsubscribe at any time.
Hulton Archive / Getty Images We warn readers that the content of this study is confronting and may be distressing. In April 1789, the first smallpox epidemic among Aboriginal people in the Sydney region began, just 16 months after Europeans arrived on the First Fleet. Little is known of the true scale of the epidemic,…
Just over two years ago I reported the ‘likely’ eradication of feral pigs from Australia’s third-largest (4,405 km2) island — Kangaroo Island. I indicated ‘likely’ because the program still required the proof-of-eradication phase to be completed before an official declaration could be made. Yesterday I had the immense honour to take part in the official…
Have you ever done any research that relied to any degree on Indigenous Knowledges? How did you cite those Knowledges, if at all? It’s probably time we rethink how we engage with Indigenous Knowledge systems. In a new article published in BioScience, we — a large group of Indigenous and non-Indigenous scholars in Australia —…