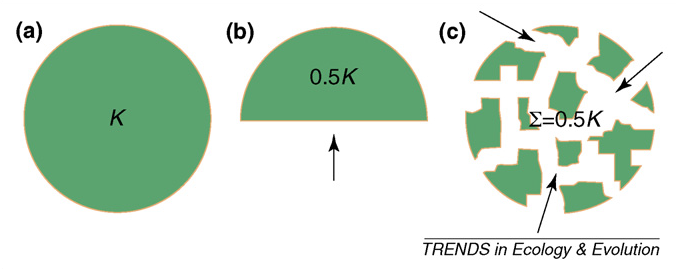
Figure 2 from Brook et al. (2008): Synergies among threatening processes relative to habitat loss and fragmentation. a) A large population within unmodified, contiguous habitat occupies all available niches so that long-term abundance fluctuates near full carrying capacity (K). b) When habitat is reduced (e.g., 50 % area loss), total abundance declines accordingly. c) However, this simple habitat-abundance relationship is complicated by the spatial configuration of habitat loss. In this example, all remaining fragmented subpopulations might fall below their minimum viable population (MVP) sizes even though total abundance is the same proportion of K as in panel B. As such, limited connectivity between subpopulations implies much greater extinction risk than that predicted for the same habitat loss in less fragmented landscapes. Further synergies (positive feedbacks among threatening processes; black arrows) might accompany high fragmentation, such as enhanced penetration of predators, invasive species or wildfire, micro-habitat edge effects, and reduced resistance to drought with climate change.
This is, perhaps, one of the most important concepts that the field of conservation biology has identified as a major driver of extinction. It may appear on the surface a rather simple notion that the more ‘habitat’ you remove, the fewer species (and individuals) there will be (see MacArthur & Wilson’s Classic contribution: The Theory of Island Biogeography), but it took us decades (yes, embarrassingly – decades) to work out that fragmentation is bad (very, very bad).
Habitat fragmentation occurs when a large expanse of a particular, broadly defined habitat ‘type’ is reduced to smaller patches that are isolated by surrounding, but different habitats. The surrounding habitat is typically defined a ‘matrix’, and in the case of forest fragmentation, generally means ‘degraded’ habitat (fewer native species, urban/rural/agricultural development, etc.).
Fragmentation is bad for many reasons: it (1) reduces patch area, (2) increases isolation among populations associated with fragments, and (3) creates ‘edges’ where unmodified habitat abuts matrix habitat. Each of these has dire implications for species, for we now know that (1) the smaller an area, the fewer individuals and species in can contain, (2) the more isolated a population, the less chance immigrants will ‘rescue’ it from catastrophes, and (3) edges allow the invasion of alien species, make the microclimate intolerable, increase access to bad humans and lead to cascading ecological events (e.g., fire penetration). Make no mistake, the more fragmented an environment, the worse will be the extinction rates of species therein.
What’s particularly sad about all this is that fragmentation was actually seen as a potentially GOOD thing by conservation biologists for many long years. The so-called SLOSS (Single Large or Several Small) debate pervaded the early days of conservation literature. The debate was basically the argument that several small reserves would provide more types of habitat juxtapositions and more different species complexes, making overall diversity (species richness) higher, than one large reserve. It was an interesting, if not deluded, intellectual debate because both sides presented some rather clever theoretical and empirical arguments. Part of the attraction of the ‘Several Small’ idea was that it was generally easier to find series of small habitat fragments to preserve than one giant no-go area.
However, we now know that the ‘Several Small’ idea is completely inferior because of the myriad synergistic effects of fragmentation. It actually took Bruce Wilcox and Dennis Murphy until 1985 to bring this to everyone’s attention in their classic paper The effects of fragmentation on extinction to show how silly the SLOSS debate really was. It wasn’t, however, until the mid- to late 1990s that people finally started to accept the idea that fragmentation really was one of the biggest conservation evils. Subsequent work (that I’ll showcase soon on ConservationBytes.com) finally put the nail in the SLOSS debate coffin, and indeed, we haven’t heard a whisper of it for over a decade.
For more general information, I invite you to read the third chapter in our book Tropical Conservation Biology entitled Broken homes: tropical biotas in fragmented landscapes, and our recent paper in Trends in Ecology and Evolution entitled Synergies among extinction drivers under global change.
CJA Bradshaw










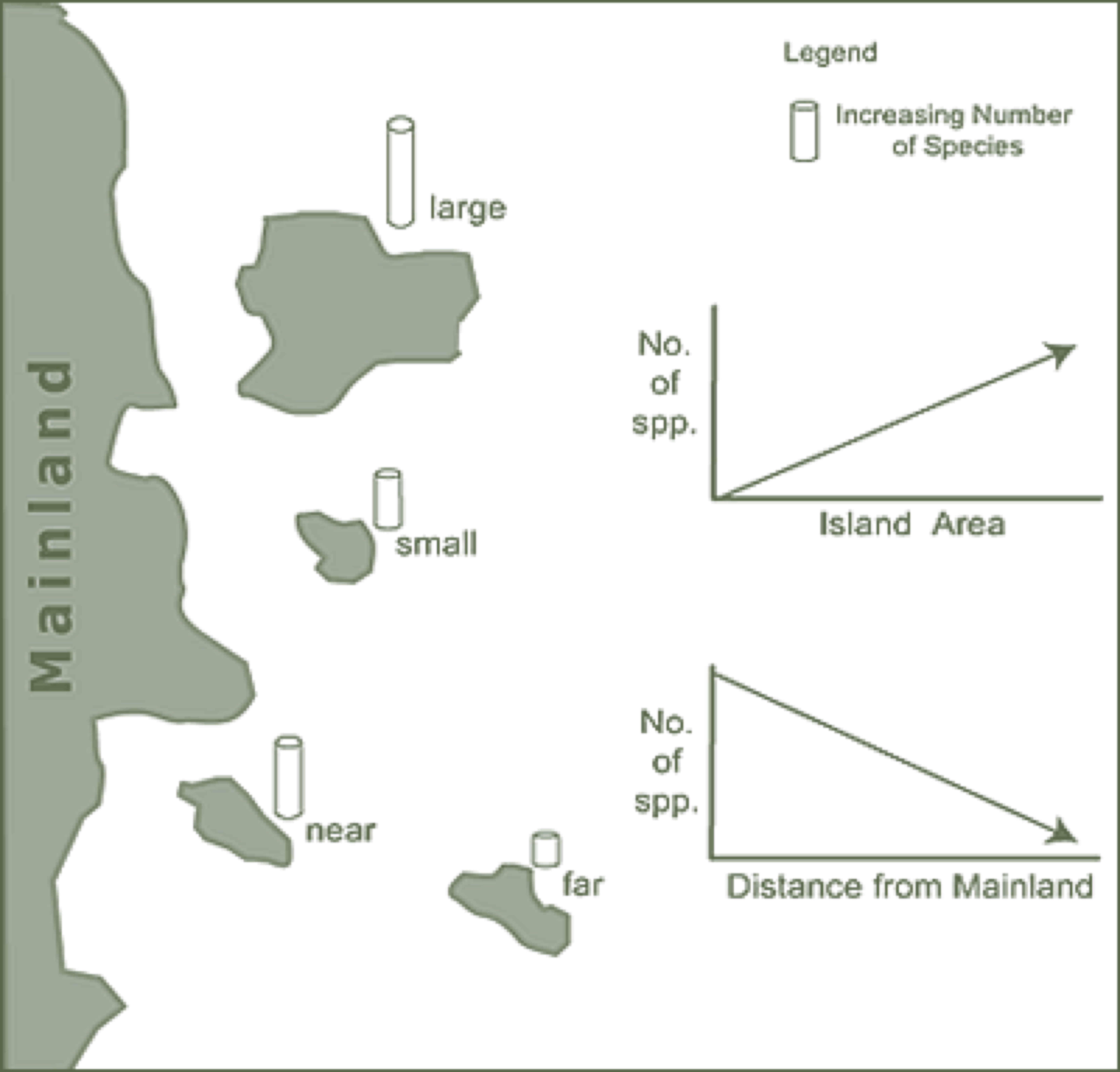 I’ve just read an elegant little study that has identified the main determinants of differences in the slope of species-area curves and species-accumulation curves.
I’ve just read an elegant little study that has identified the main determinants of differences in the slope of species-area curves and species-accumulation curves.