Have you ever watched a nature documentary and marvelled at the intricate dance of life unfolding on screen? From the smallest insect to the largest predator, every creature plays a role in the grand performance of our planet’s biosphere. But what happens when one of these performers disappears?
In this post, we delve into our recent article Estimating co-extinction risks in terrestrial ecosystems just published in Global Change Biology, in which we discuss the cascading effects of species loss and the risks of ‘co-extinction’.
But what does ‘co-extinction’ really mean?
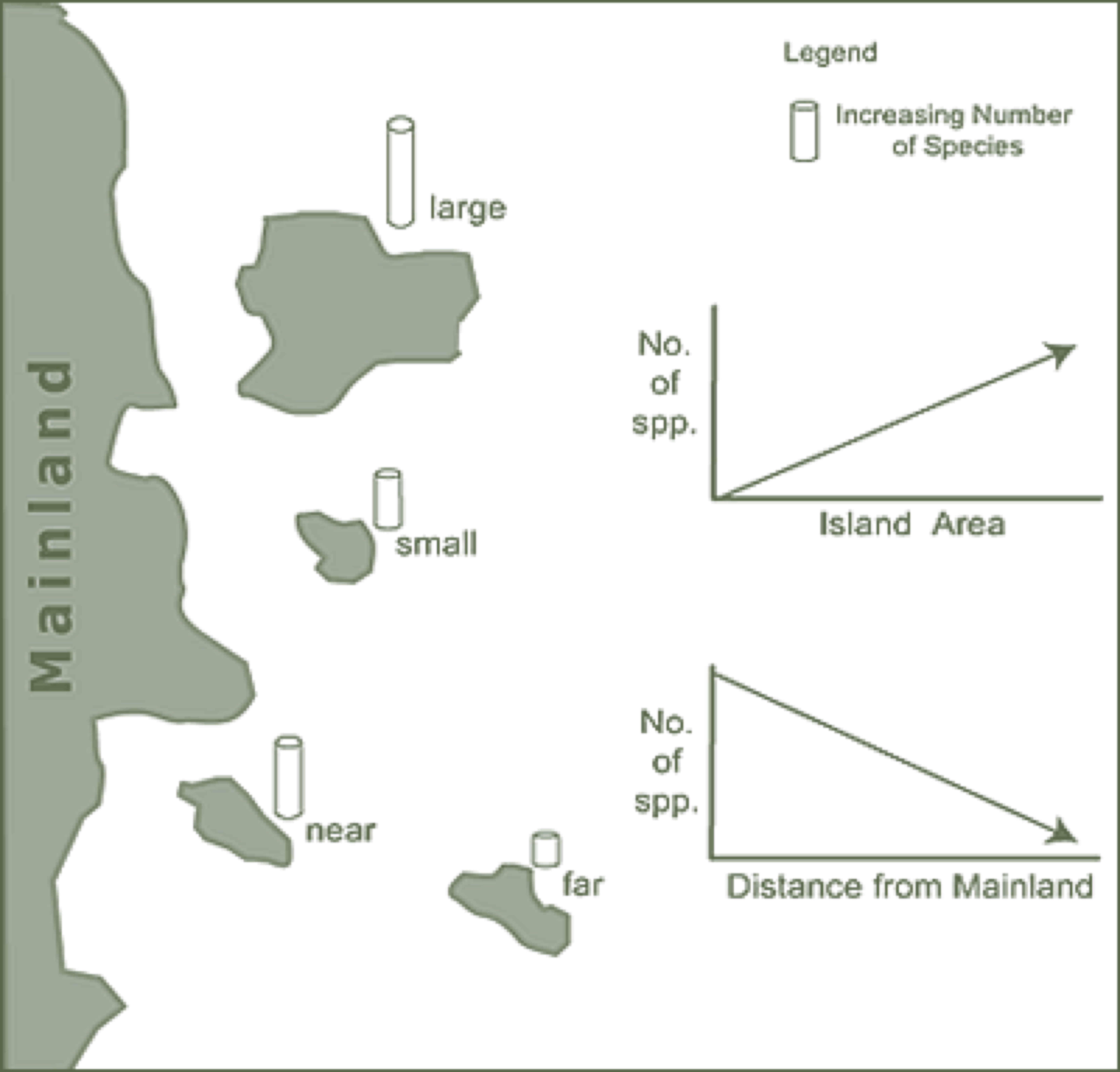
Imagine an ecosystem as a giant web of interconnected species. Each thread represents a relationship between two species — for example, a bird that eats a certain type of insect, or a plant that relies on a specific species of bee for pollination. Now, what happens if one of these species in the pair disappears? The thread breaks and the remaining species loses an interaction. This could potentially lead to its co-extinction, which is essentially the domino effect of multiple species losses in an ecosystem.
A famous example of this effect can be seen with the invasion of the cane toad (Rhinella marina) across mainland Australia, which have caused trophic cascades and species compositional changes in these communities.

Read the rest of this entry »
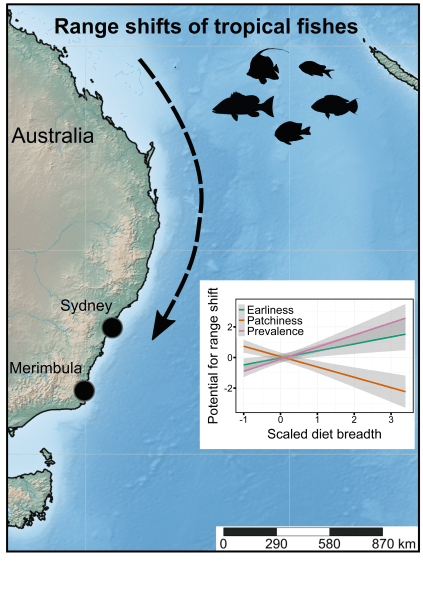 Ecologists often rely on measuring certain elements of a species’ characteristics, behaviour, or morphology to determine if these — what we call ‘traits’ — give them certain capacities to exploit their natural environments. While sometimes a bit arbitrarily defined, the traits that can be measured are many indeed, and sometimes they reveal rather interesting elements of a species’ resilience in the face of environmental change.
Ecologists often rely on measuring certain elements of a species’ characteristics, behaviour, or morphology to determine if these — what we call ‘traits’ — give them certain capacities to exploit their natural environments. While sometimes a bit arbitrarily defined, the traits that can be measured are many indeed, and sometimes they reveal rather interesting elements of a species’ resilience in the face of environmental change.




 This might seem a little left-of-centre for
This might seem a little left-of-centre for