Coral reefs are much more than just a pretty place to visit. They are among the world’s richest ecosystems, hosting about a third of all marine species.
These reefs also directly benefit more than a billion people, providing livelihoods and food security, as well as protection from storms and coastal erosion.
Without coral reefs, the world would be a much poorer place. So when corals die or become damaged, many people try to restore them. But the enormity of the task is growing as the climate keeps warming.
In our new research, we examined the full extent of existing coral restoration projects worldwide. We looked at what drives their success or failure, and how much it would actually cost to restore what’s already been lost. Restoring the reefs we’ve already lost around the world could cost up to A$26 trillion.
Bleached Acropora corals in the Maldives.Davide Seveso/University of Milan
When sea temperatures climb above the seasonal average for sustained periods, corals can become bleached. They lose colour as they expel their symbiotic algae when stressed, revealing the white skeleton underneath. Severe bleaching can kill coral.
Following my annual tradition, I present the retrospective list of the ‘top’ 20 influential papers of 2022 as assessed by experts in Faculty Opinions(formerly known as F1000). These are in no particular order. See previous years’ lists here: 2021, 2020, 2019, 2018, 2017, 2016, 2015, 2014, and 2013.
Now that the Australian election has been called for next month, here are a few cartoon reminders of the state of environmental politics in this country (hint: they’re abysmal). I’ve surpassed my normal 6 cartoons/post here in this second set for 2022 because, well, our lives depend on the outcome of 21 May. See full stock of previous ‘Cartoon guide to biodiversity loss’ compendia here.
Following my annual tradition, I present the retrospective list of the ‘top’ 20 influential papers of 2021 as assessed by experts in Faculty Opinions(formerly known as F1000). These are in no particular order. See previous years’ lists here: 2020, 2019, 2018, 2017, 2016, 2015, 2014, and 2013.
The intensity of threats to biodiversity from human endeavour becomes weaker as the distance to them increases.
As you move away from the big city to enjoy the countryside, you’ll notice the obvious increase in biodiversity. Even the data strongly support this otherwise subjective perception — there is a positive correlation between the degree we destroy habitat, harvest species, and pollute the environment, and the distance from big cities.
Remote locations are therefore usually considered safe havens and potential reservoirs for biodiversity. But our new study published recently in Nature Communications shows how this obvious pattern depicts only half of the story, and that global conservation management and actions might benefit from learning more about the missing part.
Communities are not just lists of individual species. Instead, they consist of complex networks of ecological interactions linking interdependent species. The structure of such networks is a fundamental determinant of biodiversity emergence and maintenance. However, it also plays an essential role in the processes of biodiversity loss. The decline or disappearance of some species might have detrimental —often fatal — effects on their associates. For example, a parasite cannot survive without its hosts, as much as a predator will starve without prey, or a plant will not reproduce without pollinators.
Events where a species disappears following the loss of other species on which it depends are known as co-extinctions, and they are now recognised as a primary driver of the ongoing global biodiversity crisis. The potential risk stemming from ecological dependencies is a major concern for all ecological systems.
It is possible to cultivate corals in the sea like growing a nursery of trees to restore a burned forest. Cultivated corals grow faster than wild corals and can be outplanted to increase the healthy area of damaged reefs. Incorporated in projects of citizen science and ecotourism, this activity promotes environmental awareness about coral reefs, the marine ecosystem that is both the most biodiverse and the most threatened by global change.
When I finished by undergraduate studies in the 1980s, I met several top Spanish marine biologists to prospect my first job ever in academia. In all one-to-one interviews I had, I was asked what my interests were. And when I described that I wanted to study ways of modifying impacted marine ecosystems to restore their biodiversity, a well-known professor judged that my proposition was an inviable form of jardinería marina (marine gardening) ― those words made me feel embarrassed and have remained vivid in my professional imagination since. Neither the expert nor the young researcher knew at the time that we were actually talking about ecological restoration, a discipline that was being formalised exactly then by botanists in their pledge to recover pre-European conditions for North American grasslands (1).
Aspects of coral gardening. The photos show (top) a diver scraping off (with the aid of a toothbrush) algae, sponges and parasites that compete for light and nutrients with the coral fragments under cultivation along suspended ropes (Cousin Island, Seychelles), (middle) coral outplantings in the Gulf of Eliat (Red Sea) hosting a diverse community of fish that clean off the biofouling for free (21), and (bottom) a donor colony farmed off Onna (Okinawa, Japan) (12). Photos courtesy of Luca Saponari (Cousin), Buki Rinkevich (Eliat) and Yoshimi Higa / Onna Village Fishery Cooperative.
Today, the term coral gardening encompasses the suite of methods to cultivate corals (tiny colonial jellyfish with an external skeleton and a carnivorous diet) and to outplant them into the wild to boost the growth of coral reefs following perturbations (2). In the face of the decline of coral reefs globally, due to the combination of climate change, pollution, and overfishing (3), this type of mariculture has gathered momentum in the last three decades and is currently being applied to more than 100 coral species in all the main reefs of our seas and oceans (4-6).
When snorkelling in a reef, it’s natural to think of coral colonies as a colourful scenography where fish act in a play. But what would happen to the fish if the stage went suddenly empty, as in Peter Brook’s 1971 Midsummer Night’s Dream? Would the fish still be there acting their roles without a backdrop?
This question is not novel in coral-reef science. Ecologists have often compared reef fish diversity and biomass in selected localities before and after severe events of coral mortality. Even a temporary disappearance of corals might have substantial effects on fish communities, sometimes resulting in a local disappearance of more than half of local fish species.
Considering the multiple, complex ways fish interact with — and depend on — corals, this might appear as an obvious outcome. Still, such complexity of interactions makes it difficult to predict how the loss of corals might affect fish diversity in specific contexts, let alone at the global scale.
Focusing on species-specific fish-coral associations reveals an inconsistent picture with local-scale empirical observations. When looking at the fraction of local fish diversity that strictly depends on corals for food and other more generic habitat requirements (such as shelter and reproduction), the global picture suggests that most fish diversity in reef locality might persist in the absence of corals.
The mismatch between this result and the empirical evidence of a stronger coral dependence suggests the existence of many hidden ecological paths connecting fish to corals, and that those paths might entrap many fish species for which the association to corals is not apparent.
“I wish it need not have happened in my time,” said Frodo. “So do I,’ said Gandalf, “and so do all who live to see such times. But that is not for them to decide. All we have to decide is what to do with the time that is given us.”
Frodo Baggins and Gandalf, The Fellowship of the Ring
Today, 16 high-profile scientists and I published what I describe as a ‘cold shower’ about society’s capacity to avoid a ghastly future of warfare, disease, inequality, persecution, extinction, and suffering.
And it goes way beyond just the plight of biodiversity.
No one who knows me well would mistake me for an optimist, try as I might to use my colleagues’ and my research for good. Instead, I like to describe myself as a ‘realist’. However, this latest paper has made even my gloomier past outputs look downright hopeful.
And before being accused of sensationalism, let me make one thing abundantly clear — I sincerely hope that what we describe in this paper does not come to pass. Not even I am that masochistic.
I am also supportive of every attempt to make the world a better place, to sing about our successes, regroup effectively from our failures, and maintain hope in spite of evidence to the contrary.
But failing to acknowledge the magnitude and the gravity of the problems facing us is not just naïve, it is positively dangerous and potentially fatal.
In the first of two consecutive interviews with climate-change experts, authors, editors and readers of the Spanish magazine Quercus have a chat with Ken Caldeira, a global-ecology researcher at the Carnegie Institution for Science (Washington, USA). His responses attest that the climate system is complex, and that we need to be practical in dealing with the planet’s ongoing climate emergency.
PhD in atmospheric sciences and professor at Stanford University (USA), Ken Caldeira has pioneered the study of ocean acidification and its impact on coral reefs (1) and geoengineering solutions to mitigate anthropogenic climate change by extracting carbon from the atmosphere and reflecting solar radiation (2, 3). He has also been part of the Intergovernmental Panel on Climate change (IPCC) and assessed zero-emissions scenarios (4, 5). To the right, Ken manoeuvers a drone while collecting aerial data from the Great Barrier Reef in Australia (6). Source.
SARS-Covid-19 is impacting the world. In our home country, Spain, scientists argue that (i) previous budget cuts in public health have weakened our capacity to tackle the pandemic (7), and (ii) the expert panels providing advice to our government should be independent of political agendas in their membership and decisions (8). Nevertheless, the Spanish national and regional governments’ data lack the periodicity, coherence, and detail to harness an effective medical response (9). Sometimes it feels as if politics partly operate by neglecting the science needed to tackle challenges such as the covid pandemic or climate change.
Having said that, even if a country has cultivated and invested in the best science possible, people have difficulties coming to terms with the idea that scientists work with probabilities of alternative scenarios. As much as there are different ways of managing a pandemic, scientists differ about how to mitigate the ecological, economic, and health impacts of a high-carbon society.
Thus, a more and more common approach is to make collective assessments (elicitations) by weighing different points of view across experts — for instance, to establish links between climate change and armed conflict (10) or to evaluate the role of nuclear energy as we transition to a low-carbon energy-production model (11). The overarching goal is to quantify consensus based on different (evidence-based) opinions.
The questions we here ask Ken Caldeira could well have different answers if asked of other experts. Still, as Ken points out, it is urgent that (of the many options available) we use the immense and certainty-proof knowledge we have already about climate change to take actions that work.
Interview done 23 January 2020
We italicise each question and the name of the person asking the question and cite one to three relevant publications per question. For expanding on Ken Caldeira’s views on climate change, see a sample of his public talks here and here and newspaper articles here and here.
The Australian awakening to its environmental dilemmas was a little more sluggish than elsewhere in the New World. Not only did Europeans arrive in Australia en masse only about 250 years ago, they had a more limited view of their new landscape, and were, at least initially, constrained by the harshness of their new home. Those mostly British settlers brought with them the fully formed ideas of development and progress shaped by centuries of land use in the Motherland. That ideal of conquering wilderness and transforming it into the bucolic landscape typical of the English countryside was their driving force.
The early settlers viewed the Australian bush as ugly and monotonous, features that could only be overcome by human occupation and cultivation. This neo-classical view, homesickness and the Romantic desire to transform their homes and farms into an image of those from their homeland, were defining forces in early Australian history. Unlike in Europe, though, where there were cultural taboos associated with forest degradation — bound in mysticism, spirituality, folklore and politics — no such restrictions applied to the unfamiliar Australian bush.
In fact, the Australian government passed the Crown Lands Alienation Act in 1861, which was designed to ‘open up’ the colony to settlement, and penalized landholders for not clearing the land (via a forfeit of the land back to the Crown). That single Act guaranteed the deforestation wave would continue for over a 100 years. That, and the persistent desire to make the new land look as much as possible as the old, has ensured that continuing demise of Australia’s biodiversity.
Clearing for agriculture in early settlement. Anonymous, Government Farm at Castle Hill, circa 1803. Watercolour, 24×35 cm. Permission to reproduce courtesy of the Mitchell Library, State Library of New South Wales
Interestingly, clashes over land use between the settlers and Indigenous peoples were probably some of the first demonstrations of what today we would call ‘environmentalism’ in Australia. Aboriginal nations were intent on preserving their way of life (and indeed, their lives) in the face of the settlers’ onslaught. But this was seen, at most, as a mild inconvenience for the new Australians who in response invoked the idea of terra nullius — that no one owned the land, making it available to anyone (white) who wished to ‘improve’ (clear) it. Read the rest of this entry »
Ecologists often rely on measuring certain elements of a species’ characteristics, behaviour, or morphology to determine if these — what we call ‘traits’ — give them certain capacities to exploit their natural environments. While sometimes a bit arbitrarily defined, the traits that can be measured are many indeed, and sometimes they reveal rather interesting elements of a species’ resilience in the face of environmental change.
As we know, climate change is changing the way species are distributed around the planet, for the main (and highly simplified) reason that the environments in which they’ve evolved and to which they have adapted are changing.
In the simplest case, a warming climate means that there is a higher and higher chance you’ll experience temperatures that really don’t suit you that well (think of a koala or a flying fox baking in a tree when the thermometer reads +45° in the shade). Just like you seeking those nice, air-conditioned spaces on a scorcher of a day, species like to move to where conditions are more acceptable to their particular physiologies and behaviours.
Ecologists use life-history traits to predict which species have the highest probability of moving to new areas in response to climate change. Most studies into this phenomenon have largely ignored that range shifts in fact occur in sequential stages: (1) the species arrives in a new place for the first time, (2) its population increases in size (and extent), and (3) it can continue to persist in the new spot. Read the rest of this entry »
I often find myself in a position explaining to non-professionals just how bad the state of global biodiversity really is. It turns out too that even quite a few ecologists seem to lack an appreciation of the sheer magnitude of damage we’ve done to the planet.
The loss of biodiversity that has occurred over the course of our species’ time on Earth is staggering. This loss is now truly planetary in scale and caused by human actions, albeit the severity of which is unequally distributed across the globe1. While Sandra Díaz and company recently summarised the the extent of the biodiversity crisis unfolding1 well in their recent synopsis of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services (IPBES)2 report, I’m going to repeat some of the salient summary statements here, and add a few others. Read the rest of this entry »
For the last five years I’ve published a retrospective list of the ‘top’ 20 influential papers of the year as assessed by experts in F1000 Prime — so, I’m doing so again for 2018 (interesting side note: six of the twenty papers highlighted here for 2018 appear in Science magazine). See previous years’ posts here: 2017, 2016, 2015, 2014, and 2013.
Female green turtles (Chelonia mydas) spawning (top) and diving (bottom) on Raine Island (Great Barrier Reef, Queensland, Australia) — photos courtesy of Ian Bell. This species is ‘Endangered’ globally since 1982, mainly from egg harvesting (poaching conflict in Mexico for olive ridleyLepidochelys olivacea featured by National Geographic’s video here), despite the success of conservation projects (39). Green turtles inhabit tropical and subtropical seas in all oceans. Adults can grow > 150 kg and live for up to ~ 75 years. Right after birth, juveniles venture into the open sea to recruit ultimately in coastal areas until sexual maturity. They then make their first reproductive migration, often over 1000s of km (see footage of a real dive of a camera-equipped green turtle), to reach their native sandy beaches where pregnant females will lay their eggs. Each female can deposit more than one hundred eggs in her nest, and in several clutches in the same season because they can store the sperm from multiple mating events.
When sex is determined by the thermal environment, males or females might predominate under sustained climatic conditions. A study about marine turtles from the Great Barrier Reef illustrates how feminisation of a population can be partitioned geographically when different reproductive colonies are exposed to contrasting temperatures.
Fortunately, most people in Western societies already perceive that we live in a complex blend of sexual identities, far beyond the kind of genitals we are born with. Those identities start to establish themselves in the embryo before the sixth week of pregnancy. In the commonest scenario, for a human foetusXY with one maternal chromosome (X) and one paternal (Y) chromosome, the activation of the Sry gen (unique to Y) will trigger the differentiation of testicles and, via hormonal pathways, the full set of male characteristics (1).
Absence of that gene in an XX embryo will normally lead to a woman. However, in just one of many exceptions to the rule, Sry-expression failure in XY individuals can result in sterile men or ambiguous genitals — along a full gradient of intermediate sexes and, potentially, gender identities. A 2015 Nature ‘News’ feature echoes two extraordinary cases: (i) a father of four children found to bear a womb during an hernia operation, and (ii) a pregnant mother found to host both XX and XY cells during a genetic test – with her clinical geneticist stating “… that’s the kind of science-fiction material for someone who just came in for an amniocentesis” (2). These real-life stories simply reflect that sex determination is a complex phenomenon.
As I have done for the last four years (2016, 2015, 2014, 2013), here’s another retrospective list of the top 20 influential conservation papers of 2017 as assessed by experts in F1000 Prime.
Hydrozoan polyps living on the surface of a coral (photo credit: S. Montano)
Corals could have some unexpected allies to cope with the multi-faceted threats posed by climate change.
In a new study published today in Proceedings of the Royal Society B, Montano and colleagues show how tiny hydrozoans smaller than 1 mm and commonly found in dense colonies on the surface of hard corals (see above photo) play an important ecological role.
Visually examining ~ 2500 coral colonies in both Maldivian and Saudi Arabian reefs, the scientists searched for signs of predation, temperature-induced stress, and disease. For each colony, they also recorded the presence of symbiotic hydrozoans. They demonstrated that corals living in association with hydrozoans are much less prone to be eaten by corallivorous (i.e., ‘coral-eating’) fish and gastropods than hydrozoan-free corals.
A likely explanation for this pattern could be the deterring action of hydrozoan nematocysts (cells capable of ejecting a venomous organelle, which are the same kinds found in jellyfish tentacles). An individual hydrozoan polyp of less than 1 mm clearly cannot cope with a corallivorous fish that is a billions of times larger, yet hydrozoans can grow at high densities on the surface of corals (sometimes > 50 individuals per cm2). This creates a sort of a continuous, ‘urticating‘ carpet that can discourage fish from foraging. Read the rest of this entry »
Three of the coral species studied by Muir (2): (a) Acropora pichoni: Pohnpei Island, Pacific Ocean — deep-water species/IUCN ‘Near threatened’; (b) Acropora divaricate: Maldives, Indian ocean — mid-water species/IUCN ‘Near threatened’; and (c) Acropora gemmifera: Orpheus Island, Australia — shallow-water species/IUCN ‘Least Concern’. The IUCN states that the 3 species are vulnerable to climate change (acidification, temperature extremes) and demographic booms of the invading predator, the crown-of-thorns starfishAcanthaster planci. Photos courtesy of Paul Muir.
Global warming of the atmosphere and the oceans is modifying the distribution of many plants and animals. However, marine species are bound to face non-thermal barriers that might preclude their dispersal over wide stretches of the sea. Sunlight is one of those invisible obstacles for corals from the Indian and Pacific Oceans.
If we were offered a sumptuous job overseas, our professional success in an unknown place could be limited by factors like cultural or linguistic differences that have nothing to do with our work experience or expertise. If we translate this situation into biodiversity terms, one of the best-documented effects of global warming is the gradual dispersal of species tracking their native temperatures from the tropics to the poles (1). However, as dispersal progresses, many species encounter environmental barriers that are not physical (e.g., a high mountain or a wide river), and whose magnitude could be unrelated to ambient temperatures. Such invisible obstacles can prevent the establishment of pioneer populations away from the source.
Corals are ideal organisms to study this phenomenon because their life cycle is tightly geared to multiple environmental drivers (see ReefBase: Global Information System for Coral Reefs). Indeed, the growth of a coral’s exoskeleton relies on symbiotic zooxanthellae (see video and presentation), a kind of microscopic algae (Dinoflagellata) whose photosynthetic activity is regulated by sea temperature, photoperiod and dissolved calcium in the form of aragonite, among other factors.
Coral reef fishes are wonderfully diverse in size, form, and function, as well as their need for different habitats throughout the life cycle. Some species spend all of their life in the same kind of coral habitat, while others need different places to breed and feed.
Fishes requiring different habitats as they progress through life often have what we call ‘nurseries’ in which adults lay eggs and the subsequent juveniles remain, and these places are often dominated by mangroves or seagrasses (i.e., they are not part of the coral reef).
While we’ve known for quite some time that when these nursery habitats are not around, adjacent coral reefs have few, if any, of these nursery-dependent species. What we haven’t known until now is just how far the influence of nurseries extends along a coral reef.
In other words, if a nursery is present, just how many new recruits do different areas of a reef receive from it? Read the rest of this entry »
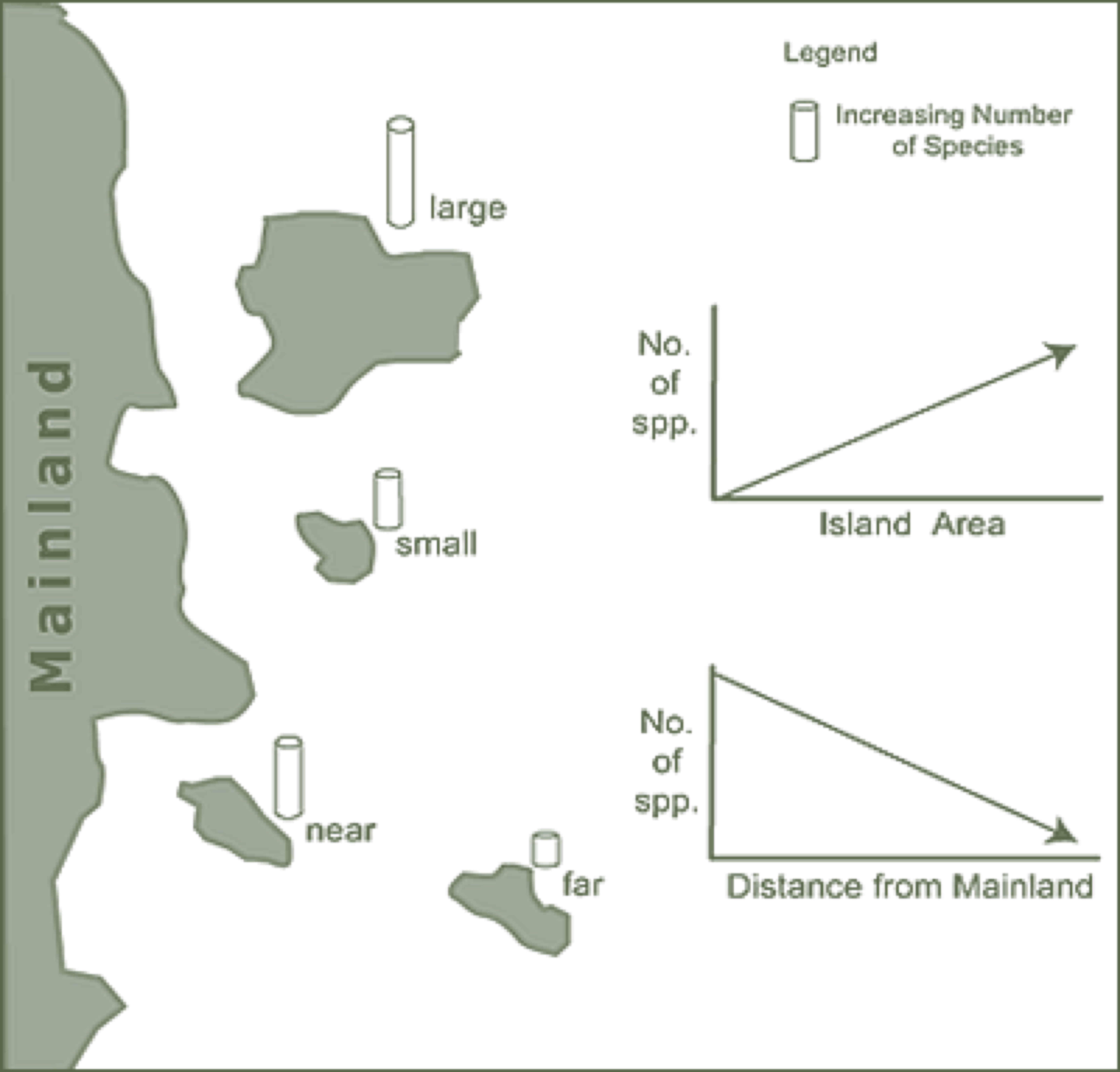
I’ve just read an elegant little study that has identified the main determinants of differences in the slope of species-area curves and species-accumulation curves.
That’s a bit of a mouthful for the uninitiated, so if you don’t know much about species-area theory, let me give you a bit of background for why this is an important new discovery.
Perhaps one of the only ‘laws’ in ecology comes from the observation that as you sample from larger and larger areas of any habitat type, the number of species tends to increase. This of course originates from MacArthur & Wilson’s classic book, The Theory of Island Biography (1967), and while simple in basic concept, it has since developed into a multi-headed Hydra of methods, analysis, theory and jargon.
One of the most controversial aspects of generic species-area relationships is the effect of different sampling regimes, a problem I’ve blogged about before. Whether you are sampling once-contiguous forest of habitat patches in a ‘matrix’ of degraded landscape, a wetland complex, a coral reef, or an archipelago of true oceanic islands, the ‘ideal’ models and the interpretation thereof will likely differ, and in sometimes rather important ways from a predictive and/or applied perspective. Read the rest of this entry »
While there are a lot of different combinations of traits that make some species more vulnerable to extinction than others (see examples for legumes, amphibians, sharks & teleosts, and mammals), one of the main ones is species size.
Generally speaking, larger species tend to produce fewer offspring and breed later in life than smaller species. This means that despite larger species tending to live longer than their smaller counterparts, their ‘slow’ reproductive output means that they are generally more susceptible to rapid environmental change (mainly via human intervention). In other words, their capacity for self-replacement is often too low to counteract the offtake from direct exploitation or habitat loss.
Despite a reasonable scientific understanding of this extinction-risk principle, the degree to which human disturbance affects species’ distributions is much less well quantified, and this is especially true for marine species.
With the world’s largest combined dataset of coral reef fish surveys for the entire Indo-Pacific (including the coral reef fish biodiversity hotspot — the Coral Triangle), we examined which conditions best described the distribution of fishes over a range of body sizes. Read the rest of this entry »
Get Conservation Bytes by Email Enter your email address to receive new posts on conservation, biodiversity, and environmental science directly in your inbox. No need to visit the website regularly, and you can unsubscribe at any time.
Hulton Archive / Getty Images We warn readers that the content of this study is confronting and may be distressing. In April 1789, the first smallpox epidemic among Aboriginal people in the Sydney region began, just 16 months after Europeans arrived on the First Fleet. Little is known of the true scale of the epidemic,…
Just over two years ago I reported the ‘likely’ eradication of feral pigs from Australia’s third-largest (4,405 km2) island — Kangaroo Island. I indicated ‘likely’ because the program still required the proof-of-eradication phase to be completed before an official declaration could be made. Yesterday I had the immense honour to take part in the official…
Have you ever done any research that relied to any degree on Indigenous Knowledges? How did you cite those Knowledges, if at all? It’s probably time we rethink how we engage with Indigenous Knowledge systems. In a new article published in BioScience, we — a large group of Indigenous and non-Indigenous scholars in Australia —…











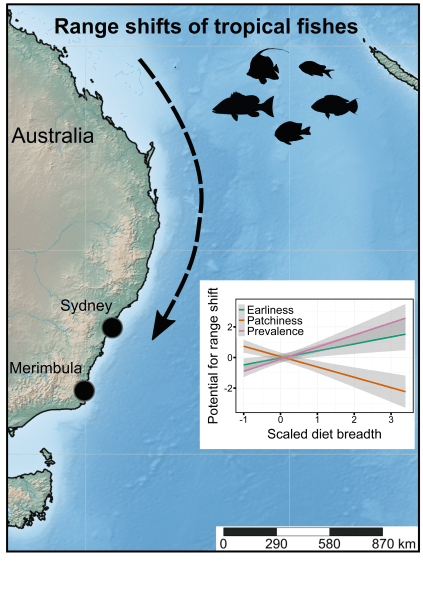 Ecologists often rely on measuring certain elements of a species’ characteristics, behaviour, or morphology to determine if these — what we call ‘traits’ — give them certain capacities to exploit their natural environments. While sometimes a bit arbitrarily defined, the traits that can be measured are many indeed, and sometimes they reveal rather interesting elements of a species’ resilience in the face of environmental change.
Ecologists often rely on measuring certain elements of a species’ characteristics, behaviour, or morphology to determine if these — what we call ‘traits’ — give them certain capacities to exploit their natural environments. While sometimes a bit arbitrarily defined, the traits that can be measured are many indeed, and sometimes they reveal rather interesting elements of a species’ resilience in the face of environmental change.