The Philippine bare-backed fruit bat (Dobsonia chapmani; body size = < 220 mm, < 150 g; IUCN status: ‘Critically Endangered A2cd’) is endemic to lowland rain forests [top habitat image] from Negros and Cebu islands. This species of flying fox had been missing from the 1970s and was declared extinct in 2002 (34). In May 2003, five specimens [one shown in the picture above] were trapped in night nets in the Calatong forest (Negros Island), a ~ 1,000-ha fragment of secondary rain forest and agricultural lands [bottom habitat image] (35). The species is reliant on fruit-bearing vegetation and caves for feeding and roosting, respectively. As with many other Philippine bats, it suffers from habitat degradation and hunting. The family Pteropodidae comprises > 150 species. Despite their Draculian look, they all feed on fruits and nectar, and act as important plant pollinators (36), as well as disease vectors such as Ebola virus (37). Flying foxes are distributed in the tropics and subtropics from the Eastern Mediterranean, through the Arabian Peninsula, Asia, Australia, and many islands of the Indian Ocean. Photos courtesy of Ely L. Alcala.
Jared Diamond (1) coined the expression ‘evil quartet’ for the four main human causes of species extinctions: habitat loss/fragmentation, overkill, introduced species and extinction chains [with climate change and extinction synergies (2), the updated expression would be ‘evil sextet”]. However, one third of ‘extinct’ mammal species has been ‘found’ again. Recent studies reveal that the probability of rediscovery depends on the cause of extinction.
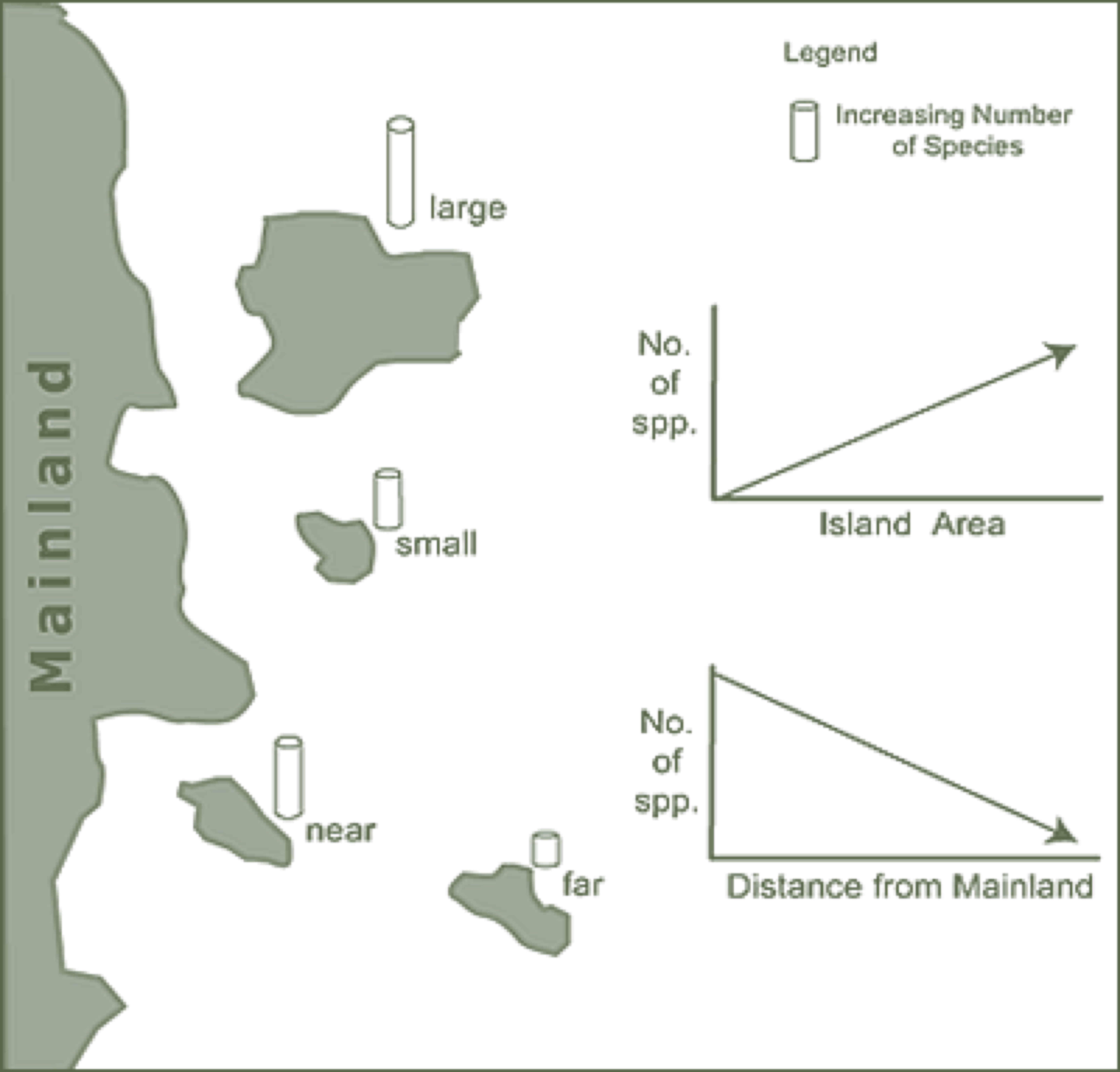
Arriving in a city to search for an old friend, I would first look in the suburb where he lived, the pub where we enjoyed a drink and some music, or the park where we used to play football. But if my friend was an outlaw, or had recently gone through a traumatic experience, my chances of finding him at his favourite spots would shrink.
If, instead of a friend, we are searching for the last survivors of an extinct-declared species, surveys also tend to take place in the habitat in which the species was previously found. Such a strategy rests on the classical hypothesis that, given the spatial distribution of a species, its gradual decline must occur from the periphery to the core of its distribution (‘range collapse’) where, in theory, the habitat should be of better quality and the number of individuals higher (3). In contrast, recent work supports that the trajectory of demise of threatened vertebrates progresses from the core to the periphery (‘range eclipse’) (4), because many perturbations make their way as a progressive wave, e.g, fire, logging or urbanisation.
Diana Fisher (5) supports the ‘range eclipse’ hypothesis for ‘extinct’ mammals which have been rediscovered. She quantifies that 60% of the new records are made from peripheral habitats, mainly when the principal cause of extirpation is habitat loss. Not only that, on average species are rediscovered at altitudes 35 % higher than historical records, and only in 5 % of the cases at the locality where it had been last seen.
Read the rest of this entry »
-34.917731
138.603034
 Last year I wrote what has become a highly viewed post here at ConservationBytes.com about the plight of the world’s freshwater biodiversity. In a word, it’s ‘buggered’.
Last year I wrote what has become a highly viewed post here at ConservationBytes.com about the plight of the world’s freshwater biodiversity. In a word, it’s ‘buggered’.