Imagine growing up beside the eastern Mediterranean Sea 14,000 years ago. You’re an accomplished sailor of the small watercraft you and your fellow villagers make, and you live off both the sea and the land.
But times have been difficult — there just isn’t the same amount of game or fish around as when you were a child. Maybe it’s time to look elsewhere for food.
Now imagine going farther than ever before in your little boat, accompanied maybe by a few others, when suddenly you spot something on the horizon. Is that an island?
The western coast of Cyprus. CJA Bradshaw / Flinders University
When you beach your boat to have a look around, you can’t believe what you’re seeing — tiny boar-sized hippos and horse-sized elephants that look like babies to your eyes. There are so many of them, and you’re hungry after the long journey.
The diminutive beasts don’t seem to show any fear. You easily kill a few and preserve the meat as best you can for the long journey back.
When you get home, you are excited to let everyone in the village know what you’ve found. Soon enough, you organise a major expedition back to the island.
Of course, we’ll never know if this kind of scenario took place, but it’s a plausible story of how and when the first humans managed to get to Cyprus. It also illustrates how they might have quickly brought about the demise of the tiny hippopotamusPhanourios minor, as well as the dwarf elephantPalaeoloxodon cypriotes.
Flooding in the Murray-Darling Basin is creating ideal breeding conditions for many native species that have evolved to take advantage of temporary flood conditions. Led by PhD candidate Rupert Mathwin, our team developed virtual models of the Murray River to reveal a crucial link between natural flooding and the extinction risk of endangered southern bell frogs (Litoria raniformis; also known as growling grass frogs).
Southern bell frogs are one of Australia’s 100 Priority Threatened Species. This endangered frog breeds during spring and summer when water levels increase in their wetlands. However, the natural flooding patterns in Australia’s largest river system have been negatively impacted by expansive river regulation that some years, sees up to 60% of river water extracted for human use.
Our latest paper describes how we built computer simulations of Murray-Darling Basin wetlands filled with simulated southern bell frogs. By changing the simulation from natural to regulated conditions, we showed that modern conditions dramatically increase the extinction risk of these beloved frogs.
The data clearly indicate that successive dry years raise the probability of local extinction, and these effects are strongest in smaller wetlands. Larger wetlands and those with more frequent inundation are less prone to these effects, although they are not immune to them entirely. The models present a warning — we have greatly modified the way the river behaves, and the modern river cannot support the long-term survival of southern bell frogs.’
In a newly announced partnership with Texas biotech company Colossal Biosciences, Australian researchers are hoping their dream to bring back the extinct thylacine is a “giant leap” closer to fruition.
Scientists at University of Melbourne’s TIGRR Lab (Thylacine Integrated Genetic Restoration Research) believe the new partnership, which brings Colossal’s expertise in CRISPR gene editing on board, could result in the first baby thylacine within a decade.
The genetic engineering firm made headlines in 2021 with the announcement of an ambitious plan to bring back something akin to the woolly mammoth, by producing elephant-mammoth hybrids or “mammophants”.
But de-extinction, as this type of research is known, is a highly controversial field. It’s often criticised for attempts at “playing God” or drawing attention away from the conservation of living species. So, should we bring back the thylacine? We asked five experts.
As someone who writes a lot of models — many for applied questions in conservation management (e.g., harvest quotas, eradication targets, minimum viable population sizes, etc.), and supervises people writing even more of them, I’ve had many different experiences with their uptake and implementation by management authorities.
Some of those experiences have involved catastrophic failures to influence any management or policy. One particularly painful memory relates to a model we wrote to assist with optimising approaches to eradicate (or at least, reduce the densities of) feral animals in Kakadu National Park. We even wrote the bloody thing in Visual Basic (horrible coding language) so people could run the module in Excel. As far as I’m aware, no one ever used it.
Others have been accepted more readily, such as a shark-harvest model, which (I think, but have no evidence to support) has been used to justify fishing quotas, and one we’ve done recently for the eradication of feral pigs on Kangaroo Island (as yet unpublished) has led directly to increased funding to the agency responsible for the programme.
According to Altmetrics (and the online tool I developed to get paper-level Altmetric information quickly), only 3 of the 16 of what I’d call my most ‘applied modelling’ papers have been cited in policy documents:
I’m pleased to announce the publication of a paper led by Kathryn Venning (KV) that was derived from her Honours work in the lab. Although she’s well into her PhD on an entirely different topic, I’m overjoyed that she persevered and saw this work to publication.
Feral cats occupy every habitat in the country, from the high tropics to the deserts, and from the mountains to the sea. They adapt to the cold just as easily as they adapt to the extreme heat, and they can eat just about anything that moves, from invertebrates to the carcases of much larger animals that they scavenge.
Cats are Australia’s bane, but you can’t help but be at least a little impressed with their resilience.
Still, we have to try our best to get rid of them where we can, or at least reduce their densities to the point where their ecological damage is limited.
Typically, the only efficient and cost-effective way to do that is via lethal control, but by using various means. These can include direct shooting, trapping, aerial poison-baiting, and a new ‘smart’ method of targeted poison delivery via a prototype device known as a Felixer™️. The latter are particularly useful for passive control in areas where ground-shooting access is difficult.
A live Felixer™️ deployed on Kangaroo Island (photo: CJA Bradshaw 2020)
A few years back the federal government committed what might seem like a sizeable amount of money to ‘eradicate’ cats from Australia. Yeah, good luck with that, although the money has been allocated to several places where cat reduction and perhaps even eradication is feasible. Namely, on islands.
For many years I’ve been interested in modelling the extinction dynamics of megafauna. Apart from co-authoring a few demographically simplified (or largely demographically free) models about how megafauna species could have gone extinct, I have never really tried to capture the full nuances of long-extinct species within a fully structured demographic framework.
That is, until now.
But how do you get the life-history data of an extinct animal that was never directly measured. Surely, things like survival, reproductive output, longevity and even environmental carrying capacity are impossible to discern, and aren’t these necessary for a stage-structured demographic model?
The answer to the first part of that question “it’s possible”, and to the second, it’s “yes”. The most important bit of information we palaeo modellers need to construct something that’s ecologically plausible for an extinct species is an estimate of body mass. Thankfully, palaeontologists are very good at estimating the mass of the things they dig up (with the associated caveats, of course). From such estimates, we can reconstruct everything from equilibrium densities, maximum rate of population growth, age at first breeding, and longevity.
But it’s more complicated than that, of course. In Australia anyway, we’re largely dealing with marsupials (and some monotremes), and they have a rather different life-history mode than most placentals. We therefore have to ‘correct’ the life-history estimates derived from living placental species. Thankfully, evolutionary biologists and ecologists have ways to do that too.
The Pleistocene kangaroo Procoptodon goliah, the largest and most heavily built of the short-faced kangaroos, was the largest and most heavily built kangaroo known. It had an unusually short, flat face and forwardly directed eyes, with a single large toe on each foot (reduced from the more normal count of four). Each forelimb had two long, clawed fingers that would have been used to bring leafy branches within reach.
So with a battery of ecological, demographic, and evolutionary tools, we can now create reasonable stochastic-demographic models for long-gone species, like wombat-like creatures as big as cars, birds more than two metres tall, and lizards more than seven metres long that once roamed the Australian continent.
Ancient clues, in the shape of fossils and archaeological evidence of varying quality scattered across Australia, have formed the basis of several hypotheses about the fate of megafauna that vanished during a peak about 42,000 years ago from the ancient continent of Sahul, comprising mainland Australia, Tasmania, New Guinea and neighbouring islands.
There is a growing consensus that multiple factors were at play, including climate change, the impact of people on the environment, and access to freshwater sources.
Just published in the open-access journal eLife, our latest CABAH paper applies these approaches to assess how susceptible different species were to extinction – and what it means for the survival of species today.
Using various characteristics such as body size, weight, lifespan, survival rate, and fertility, we (Chris Johnson, John Llewelyn, Vera Weisbecker, Giovanni Strona, Frédérik Saltré & me) created population simulation models to predict the likelihood of these species surviving under different types of environmental disturbance.
We compared the results to what we know about the timing of extinction for different megafauna species derived from dated fossil records. We expected to confirm that the most extinction-prone species were the first species to go extinct – but that wasn’t necessarily the case.
While we did find that slower-growing species with lower fertility, like the rhino-sized wombat relative Diprotodon, were generally more susceptible to extinction than more-fecund species like the marsupial ‘tiger’ thylacine, the relative susceptibility rank across species did not match the timing of their extinctions recorded in the fossil record.
Indeed, we found no clear relationship between a species’ inherent vulnerability to extinction — such as being slower and heavier and/or slower to reproduce — and the timing of its extinction in the fossil record.
In fact, we found that most of the living species used for comparison — such as short-beaked echidnas, emus, brush turkeys, and common wombats — were more susceptible on average than their now-extinct counterparts.
However, this time I’ve strayed from my recent bibliometric musings and developed something that’s more compatible with the core of my main research and interests.
Over the years I’ve taught many students the basics of population modelling, with the cohort-based approaches dominating the curriculum. Of these, the simpler ‘Leslie’ (age-classified) matrix models are both the easiest to understand and for which data can often be obtained without too many dramas.
But unless you’re willing to sit down and learn the code, they can be daunting to the novice.
Sure, there are plenty of software alternatives out there, such as Bob Lacy‘s Vortex (a free individual-based model available for PCs only), Resit Akçakaya & co’s RAMAS Metapop ($; PC only), Stéphane Legendre‘s Unified Life Models (ULM; open-source; all platforms), and Charles Todd‘s Essential (open-source; PC only) to name a few. If you’re already an avid R user and already into population modelling, you might be familiar with the population-modelling packages popdemo, OptiPopd, or sPop. I’m sure there are still other good resources out there of which I’m not aware.
But, even to install the relevant software or invoke particular packages in R takes a bit of time and learning. It’s probably safe to assume that many people find the prospect daunting.
It’s for this reason that I turned my newly acquired R Shiny skills to matrix population models so that even complete coding novices can run their own stochastic population models.
Several months ago, Daniel Blumstein of UCLA approached me with an offer — fancy leading a Special Section in a new Frontiers journal dedicated to conservation science?
I admit that my gut reaction was a visceral ‘no’, both in terms of the extra time it would require, as well as my autonomous reflex of ‘not another journal, please‘.
I had, for example, spent a good deal of blood, sweat, and tears helping to launch Conservation Letters when I acted as Senior Editor for the first 3.5 years of its existence (I can’t believe that it has been nearly a decade since I left the journal). While certainly an educational and reputational boost, I can’t claim that the experience was always a pleasant one — as has been said many times before, thefastest way to makeenemies is to become an editor.
But then Dan explained what he had in mind for Frontiers in Conservation Science, and the more I spoke with him, the more I started to think that it wasn’t a bad idea after all for me to join.
One of the most ancient peopling events of the great diaspora of anatomically modern humans out of Africa more than 50,000 years ago — human arrival in the great continent of Sahul (New Guinea, mainland Australia & Tasmania joined during periods of low sea level) — remains mysterious. The entry routes taken, whether migration was directed or accidental, and just how many people were needed to ensure population viability are shrouded by the mists of time. This prompted us to build stochastic, age-structured human population-dynamics models incorporating hunter-gatherer demographic rates and palaeoecological reconstructions of environmental carrying capacity to predict the founding population necessary to survive the initial peopling of late-Pleistocene Sahul.
—
As ecological modellers, we are often asked by other scientists to attempt to render the highly complex mechanisms of entire ecosystems tractable for virtual manipulation and hypothesis testing through the inevitable simplification that is ‘a model’. When we work with scientists studying long-since-disappeared ecosystems, the challenges multiply.
Add some multidisciplinary data and concepts into the mix, and the complexity can quickly escalate.
This is how we tackled one of these big questions: just how did the first anatomically modern Homo sapiens make it to the continent and survive?
At that time, Australia was part of the giant continent of Sahul that connected New Guinea, mainland Australia, and Tasmania at times of lower sea level. In fact, throughout most of last ~ 126,000 years (late Pleistocene and much of the Holocene), Sahul was the dominant landmass in the region (see this handy online tool for how the coastline of Sahul changed over this period).
I published this last week on The Conversation, and now reproducing it here for CB.com readers.
—
Two days ago, the last male northern white rhino (Ceratotherium simum cottoni) died. His passing leaves two surviving members of his subspecies: both females who are unable to bear calves.
Even though it might not be quite the end of the northern white rhino because of the possibility of implanting frozen embryos in their southern cousins (C. simum simum), in practical terms, it nevertheless represents the end of a long decline for the subspecies. It also raises the question: how many individuals does a species need to persist?
Fiction writers have enthusiastically embraced this question, most often in the post-apocalypsegenre. It’s a notion with a long past; the Adam and Eve myth is of course based on a single breeding pair populating the entire world, as is the case described in the Ragnarok, the final battle of the gods in Norse mythology.
This idea dovetails neatly with the image of Noah’s animals marching “two by two” into the Ark. But the science of “minimum viable populations” tells us a different story.
I love it when a good collaboration bears fruit, and our latest paper is a good demonstration of that principle.
It all started a few years ago with an ARC Linkage Project grant we received to examine how the whaler shark fishing industry in Australia might manage its stocks better.
As I’m sure many are aware, sharks around the world aren’t doing terribly well (surprise, surprise — yet another taxon suffering at the hands of humankind). And while some populations (‘stocks’, in the dissociative parlance of the fishing industry) are doing better than others, and some countries have a better track record in managing these stocks than others, the overall outlook is grim.
One of the main reasons sharks tend to fair worse than bony fishes (teleosts) for the same fishing effort is their ‘slow’ life histories. It doesn’t take an advanced quantitative ecology degree to understand that growing slowly, breeding late, and producing few offspring is a good indication that a species can’t handle too much killing before populations start to dwindle. As is the case for most large shark species, I tend to think of them in a life-history sense as similar to large terrestrial mammals.
Now, you’d figure that a taxon with intrinsic susceptibility to fishing would have heaps of good data with which managers could monitor catches and quotas so that declines could be avoided. However, the reality is generally the inverse, with many populations having poor information regarding vital rates (e.g., survival, fertility), age structure, density feedback characteristics, and even simple estimates of abundance. Without such key information, management tends to be ad hoc and often not very effective. Read the rest of this entry »
I’ve recently read perhaps the most comprehensive treatise of forest fragmentation research ever compiled, and I personally view this rather readable and succinct review by Bill Laurance and colleagues as something every ecology and conservation student should read.
The ‘Biological Dynamics of Forest Fragments Project‘ (BDFFP) is unquestionably one of the most important landscape-scale experiments ever conceived and implemented, now having run 38 years since its inception in 1979. Indeed, it was way ahead of its time.
Experimental studies in ecology are comparatively rare, namely because it is difficult, expensive, and challenging in the extreme to manipulate entire ecosystems to test specific hypotheses relating to the response of biodiversity to environmental change. Thus, we ecologists tend to rely more on mensurative designs that use existing variation in the landscape (or over time) to infer mechanisms of community change. Of course, such experiments have to be large to be meaningful, which is one reason why the 1000 km2 BDFFP has been so successful as the gold standard for determining the effects of forest fragmentation on biodiversity.
With the official start of the new ARCCentre of Excellence for Australian Biodiversity and Heritage (CABAH) in July, I am pleased to announce two new CABAH-funded postdoctoral positions (a.k.a. Research Associates) in my global ecology lab at Flinders University in Adelaide (Flinders Modelling Node).
One of these positions is a little different, and represents something of an experiment. The Research Associate in Palaeo-Vegetation Modelling is being restricted to women candidates; in other words, we’re only accepting applications from women for this one. In a quest to improve the gender balance in my lab and in universities in general, this is a step in the right direction.
The project itself is not overly prescribed, but we would like something along the following lines of inquiry: Read the rest of this entry »
You couldn’t really do ecology if you didn’t know how to construct even the most basic mathematical model — even a simple regression is a model (the non-random relationship of some variable to another). The good thing about even these simple models is that it is fairly straightforward to interpret the ‘strength’ of the relationship, in other words, how much variation in one thing can be explained by variation in another. Provided the relationship is real (not random), and provided there is at least some indirect causation implied (i.e., it is not just a spurious coincidence), then there are many simple statistics that quantify this strength — in the case of our simple regression, the coefficient of determination (R2) statistic is a usually a good approximation of this.
When you go beyond this correlative model approach and start constructing more mechanistic models that emulate ecological phenomena from the bottom-up, things get a little more complicated when it comes to quantifying the strength of relationships. Perhaps the most well-known category of such mechanistic models is the humble population viability analysis, abbreviated to PVA§.
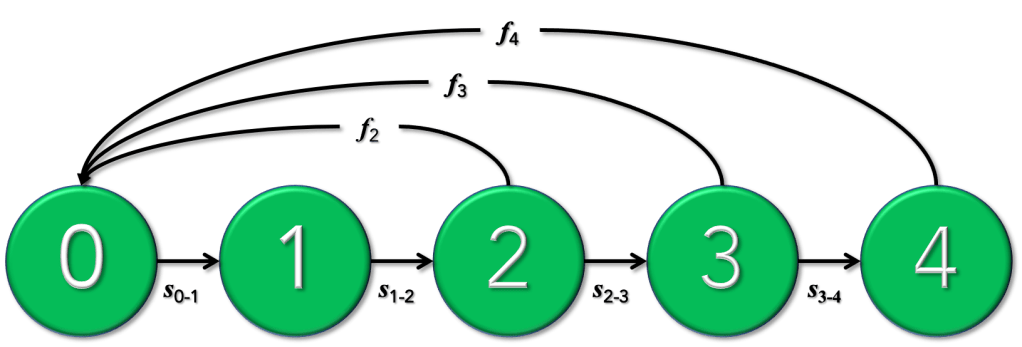
Let’s take the simple case of a four-parameter population model we could use to project population size over the next 10 years for an endangered species that we’re introducing to a new habitat. We’ll assume that we have the following information: the size of the founding (introduced) population (n), the juvenile survival rate (Sj, proportion juveniles surviving from birth to the first year), the adult survival rate (Sa, the annual rate of surviving adults to year 1 to maximum longevity), and the fertility rate of mature females (m, number of offspring born per female per reproductive cycle). Each one of these parameters has an associated uncertainty (ε) that combines both measurement error and environmental variation.
If we just took the mean value of each of these three demographic rates (survivals and fertility) and project a founding population of n = 10 individuals for 1o years into the future, we would have a single, deterministic estimate of the average outcome of introducing 10 individuals. As we already know, however, the variability, or stochasticity, is more important than the average outcome, because uncertainty in the parameter values (ε) will mean that a non-negligible number of model iterations will result in the extinction of the introduced population. This is something that most conservationists will obviously want to minimise.
So each time we run an iteration of the model, and generally for each breeding interval (most often 1 year at a time), we choose (based on some random-sampling regime) a different value for each parameter. This will give us a distribution of outcomes after the 10-year projection. Let’s say we did 1000 iterations like this; taking the number of times that the population went extinct over these iterations would provide us with an estimate of the population’s extinction probability over that interval. Of course, we would probably also vary the size of the founding population (say, between 10 and 100), to see at what point the extinction probability became acceptably low for managers (i.e., as close to zero as possible), but not unacceptably high that it would be too laborious or expensive to introduce that many individuals. Read the rest of this entry »
In July 2015 an American dentist shot and killed a male lion called ‘Cecil’ with a hunting bow and arrow, an act that sparked a storm of social media outrage. Cecil was a favourite of tourists visiting Hwange National Park in Zimbabwe, and so the allegation that he was lured out of the Park to neighbouring farmland added considerable fuel to the flames of condemnation. Several other aspects of the hunt, such as baiting close to national park boundaries, were allegedly done illegally and against the spirit and ethical norms of a managed trophy hunt.
In May 2015, a Texan legally shot a critically endangered black rhino in Namibia, which also generated considerable online ire. The backlash ensued even though the male rhino was considered ‘surplus’ to Namibia’s black rhino populations, and the US$350,000 generated from the managed hunt was to be re-invested in conservation. Together, these two incidents have triggered vociferous appeals to ban trophy hunting throughout Africa.
These highly politicized events are but a small component of a large industry in Africa worth > US$215 million per year that ‘sells’ iconic animals to (mainly foreign) hunters as a means of generating otherwise scarce funds. While to most people this might seem like an abhorrent way to generate money, we argue in a new paper that sustainable-use activities, such as trophy hunting, can be an important tool in the conservationist’s toolbox. Conserving biodiversity can be expensive, so generating money is a central preoccupation of many environmental NGOs, conservation-minded individuals, government agencies and scientists. Making money for conservation in Africa is even more challenging, and so we argue that trophy hunting should and could fill some of that gap. Read the rest of this entry »
Dick’s latest paper in Molecular Ecology is a meta-analysis designed to test whether there are any genetic grounds for NOT attempting genetic rescue for inbreeding-depressed populations. I suppose a few definitions are in order here. Genetic rescue is the process, either natural or facilitated, where inbred populations (i.e., in a conservation sense, those comprising too many individuals bonking their close relatives because the population in question is small) receive genes from another population such that their overall genetic diversity increases. In the context of conservation genetics, ‘inbreeding depression‘ simply means reduced biological fitness (fertility, survival, longevity, etc.) resulting from parents being too closely related.
Seems like an important thing to avoid, so why not attempt to facilitate gene flow among populations such that those with inbreeding depression can be ‘rescued’? In applied conservation, there are many reasons given for not attempting genetic rescue: Read the rest of this entry »
I’ve had a good week at Stanford University with Paul Ehrlich where we’ve been putting the final touches1 on our book. It’s been taking a while to put together, but we’re both pretty happy with the result, which should be published by The University of Chicago Press within the first quarter of 2015.
It has indeed been a pleasure and a privilege to work with one of the greatest thinkers of our age, and let me tell you that at 82, he’s still a force with which to be reckoned. While I won’t divulge much of our discussions here given they’ll appear soon-ish in the book, I did want to raise one subject that I think we all need to think about a little more.
The issue is what we, as ecologists (I’m including conservation scientists here), choose to study and contemplate in our professional life.
I’m just as guilty as most of the rest of you, but I argue that our discipline is caught in a rut of irrelevancy on the grander scale. We spend a lot of time refining the basics of what we essentially already know pretty well. While there will be an eternity of processes to understand, species to describe, and relationships to measure, can our discipline really afford to avoid the biggest issues while biodiversity (and our society included) are flushed down the drain?
As you might recall, Dick Frankham, Barry Brook and I recently wrote a review in Biological Conservationchallenging the status quo regarding the famous 50/500 ‘rule’ in conservation management (effective population size [Ne] = 50 to avoid inbreeding depression in the short-term, and Ne = 500 to retain the ability to evolve in perpetuity). Well, it inevitably led to some comments arising in the same journal, but we were only permitted by Biological Conservation to respond to one of them. In our opinion, the other comment was just as problematic, and only further muddied the waters, so it too required a response. In a first for me, we have therefore decided to publish our response on the arXivpre-print server as well as here on ConservationBytes.com.
—
50/500 or 100/1000 debate is not about the time frame – Reply to Rosenfeld
cite as: Frankham, R, Bradshaw CJA, Brook BW. 2014. 50/500 or 100/1000 debate is not about the time frame – Reply to Rosenfeld. arXiv: 1406.6424 [q-bio.PE]25 June 2014.
The Letter from Rosenfeld (2014) in response to Jamieson and Allendorf (2012) and Frankham et al. (2014) and related papers is misleading in places and requires clarification and correction, as follows: Read the rest of this entry »
Some of you who are familiar with my colleagues’ and my work will know that we have been investigating the minimum viable population size concept for years (see references at the end of this post). Little did I know when I started this line of scientific inquiry that it would end up creating more than a few adversaries.
It might be a philosophical perspective that people adopt when refusing to believe that there is any such thing as a ‘minimum’ number of individuals in a population required to guarantee a high (i.e., almost assured) probability of persistence. I’m not sure. For whatever reason though, there have been some fierce opponents to the concept, or any application of it.
Yet a sizeable chunk of quantitative conservation ecology develops – in various forms – population viability analyses to estimate the probability that a population (or entire species) will go extinct. When the probability is unacceptably high, then various management approaches can be employed (and modelled) to improve the population’s fate. The flip side of such an analysis is, of course, seeing at what population size the probability of extinction becomes negligible.
But most reasonable people are likely to agree that < 1 % chance of going extinct over many generations (40, in the case of our suggestion) is an acceptable target. I’d feel pretty safe personally if my own family’s probability of surviving was > 99 % over the next 40 generations.
Some people, however, baulk at the notion of making generalisations in ecology (funny – I was always under the impression that was exactly what we were supposed to be doing as scientists – finding how things worked in most situations, such that the mechanisms become clearer and clearer – call me a dreamer).
So when we were attacked in several high-profile journals, it came as something of a surprise. The latest lashing came in the form of a Trends in Ecology and Evolutionarticle. We wrote a (necessarily short) response to that article, identifying its inaccuracies and contradictions, but we were unable to expand completely on the inadequacies of that article. However, I’m happy to say that now we have, and we have expanded our commentary on that paper into a broader review. Read the rest of this entry »
A few weeks ago I wrote a post about how to run the perfect scientific workshop, which most of you thought was a good set of tips (bizarrely, one person was quite upset with the message; I saved him the embarrassment of looking stupid online and refrained from publishing his comment).
As I mentioned at the end of post, the stimulus for the topic was a particularly wonderful workshop 12 of us attended at beautiful Linnaeus Estate on the northern coast of New South Wales (see Point 5 in the ‘workshop tips’ post).
I hate to say it – mainly because it deserves as little attention as possible – but the main reason is that we needed to clean up a bit of rubbish. The rubbish in question being the latest bit of excrescence growing on that accumulating heap produced by a certain team of palaeontologists promulgating their ‘it’s all about the climate or nothing’ broken record.
Get Conservation Bytes by Email Enter your email address to receive new posts on conservation, biodiversity, and environmental science directly in your inbox. No need to visit the website regularly, and you can unsubscribe at any time.
Hulton Archive / Getty Images We warn readers that the content of this study is confronting and may be distressing. In April 1789, the first smallpox epidemic among Aboriginal people in the Sydney region began, just 16 months after Europeans arrived on the First Fleet. Little is known of the true scale of the epidemic,…
Just over two years ago I reported the ‘likely’ eradication of feral pigs from Australia’s third-largest (4,405 km2) island — Kangaroo Island. I indicated ‘likely’ because the program still required the proof-of-eradication phase to be completed before an official declaration could be made. Yesterday I had the immense honour to take part in the official…
Have you ever done any research that relied to any degree on Indigenous Knowledges? How did you cite those Knowledges, if at all? It’s probably time we rethink how we engage with Indigenous Knowledge systems. In a new article published in BioScience, we — a large group of Indigenous and non-Indigenous scholars in Australia —…