Those of us living with cats share our homes with an ancestral predator, one adapted for hunting and the frequent, exclusive consumption of meat. These instincts become fully activated outside the domestic environment, where cats pose a global threat to wildlife.
Pets are family. We celebrate their arrival with the same joy as a grand homecoming, and their absence leaves a grief as deep as losing a loved one. In bonding with cats and dogs, we often attribute human abilities and emotions to them.
But beyond this affection, domestic animals still carry the instincts and genetic legacy of their wild ancestors(1, 2). My cats — Caruso, Muesli, and Plata — have been calm and loving, but they have always enjoyed a real hunt (3). When a moth comes in through a window, they seem possessed: their mouths chitter and make clicking sounds, they leap from one piece of furniture to another, and their heads snap sharply between the insect’s position and other points in the room, calculating the best spot from which to pounce on their prey. That is why when they become feral, cats and dogs integrate into food chains like any other species: they compete for ecosystem resources, hunt and are hunted, and hybridise and exchange diseases with other carnivores (4, 5).
Top: cat eating an Eurasian blue tit (Cyanistes caeruleus), a common visitor to home gardens in Nijmegen (Netherlands). Bottom, domestic cat after hunting an Eastern cottontail rabbit (Sylvilagus floridanus) in a residential neighbourhood of Stratford (Connecticut, USA). Photos courtesy of Jelger Herder (Nijmegen) and Scott Kruitbosch (Stratford). Scott is a photographer and conservationist. Near sunset on 30/09/2020, while intently observing local wildlife, he witnessed a neighbourhood cat sneak up from behind on a cottontail feeding in open grass and grab it. For years, Scott has had extremely negative interactions, both in person and online, with local residents over these issues. These exchanges have revealed that many people show little concern for wildlife or for the dangers their outdoor cats face, and believe that their cats would not, or could not, harm wildlife.
Domestic cats are highly skilled hunters, and their predatory interactions with a wide range of prey are widely documented in social media and documentaries. Some examples include cats catching: bats and birds on the wing, butterflies, chipmunks, dragons, fishes, grasshoppers, frogs, lizards, mice, owls, rabbits, seagulls, snakes, squirrels, and wallabies. See an award-winning photo depicting wildlife with fatal injuries caused by cats recorded in 2019 at a single animal hospital in the USA, and a video showing domestic cats mimicking bird calls and some cat owners explaining that their pets reject commercial cat food after experiencing the thrill of hunting real prey. The documentary Secret Life of Cats contextualises the ecological challenges posed by free-roaming cats.
Many animals avoid contact with people. In protected areas of the African savanna, mammals flee more intensely upon hearing human conversations than when they hear lions or sounds associated with hunting. This fear of humans affects how species use and move in their habitat.
Throughout our lives, we interact with hundreds of wildlife species without stopping to think about it. These interactions can be direct, such as encountering wild animals while hiking in the mountains or driving through rural areas — or more deliberate, as when we engage with wildlife for food, sport, or trade. As hunters, fishers, and collectors, we kill more than 15,000 species of vertebrates — one-third of known diversity — a range of prey 300 times greater than that of any other predator our size (1).
Now, let’s look at it from the other side. Anyone who has survived an attack or a fatal accident, they understand that the experience is remembered for a lifetime. Likewise, animals store information about threatening or harmful encounters with humans (2). For them, adjusting their behaviour in response to human presence has implications for their survival and reproduction (3, 4), which are passed down from generation to generation (5). This ability to adapt, for example, determines which individuals, populations and species coexist with us in urbanised environments (6).
Response to dangerous sounds
Liana Zanette and her team measured the flight responses of wild mammals in the Greater Kruger National Park (South Africa) when exposed to sounds that signal danger (7) [video-summary]. To do this, Zanette recorded videos of more than 4,000 visits to 21 waterholes by 18 mammal species. During each visit, a speaker attached to a tree randomly played one of five playback sounds: hunting dogs barking, gunshots, lion growls, human conversations in a calm tone and, as a control, the songs of harmless birds.
Deep-sea sharks include some of the longest-lived vertebrates known. The record holder is the Greenland shark, with a recently estimated maximum age of nearly 400 years. Their slow life cycle makes them vulnerable to fisheries.
In the Arctic, there are whales that have survived since the time of Napoleon’s Empire; in the Atlantic, there are molluscs that were contemporary with Christopher Columbus’ voyages; and in Antarctica, there are sponges born before the Holocene when humans were still an insignificant species of hunter-gatherers (see video on lifespan variation in wildlife).
Long-lived species grow slowly and reproduce at later ages (1, 2). As a result, these animals require a long time to form abundant populations and to recover from fishing-related mortality.
Among cartilaginous fish (chimaeras, rays, sharks, and skates), the risk of extinction due to overfishing is twice as high for deep-sea species compared to coastal species, because the former have longer and slower life cycles (3).
Procreating with a relative is taboo in most human societies for many reasons, but they all stem from avoiding one thing in particular — inbreeding increases the risk of genetic disorders that can seriously compromise a child’s health, life prospects, and survival.
While we all inherit potentially harmful mutations from our parents, the effects of these mutations are often partially or completed masked if we possess two alternative variants of a gene — one from each parent. However, the children of closely related parents are more likely to inherit the same copies of harmful mutations. This is known as ‘inbreeding depression’.
But inbreeding depression can happen in any species, with the risk increasing as populations become smaller. Because many species are rapidly declining in abundance and becoming isolated from one another predominantly due to habitat destruction, invasive species, and climate change, the chances of inbreeding are also increasing.
Not only are such populations more susceptible to random disturbances, they are also victim of reduced population growth rates arising from inbreeding depression. This produces what is generally known as the ‘extinction vortex‘ — the smaller your population, the more you inbreed and produce sub-optimal offspring, leading to even more population decline and eventually extinction.
One emergency intervention that can ‘rescue’ such inbred populations from extinction (at least in the short term) is to introduce unrelated individuals from other populations in an attempt to increase genetic diversity, and therefore, the rate of population growth. While somewhat controversial because some fear introducing diseases or eroding local-area specialisation (so-called ‘outbreeding depression’), the risk-benefit ratio of this ‘genetic rescue’ is now widely considered to be worth it.
In boreal forests, many hares adopt white winter coats before the snow arrives. In a snowless landscape, these white hares lack camouflage against predators. However, their early moult from brown into white fur can increase their survival and offers an advantage as the snow season becomes progressively shorter with climate change.
Throughout the year, we wear different clothing to protect ourselves from the cold or heat and for aesthetic reasons depending on the occasion. Likewise, many animals change the colour, thickness and structure of their fur and feathers in tune with the seasons.
Snowshoe hare (Lepus americanus) in a snowy (Kluane Lake/Yukon, Canada) and snowless habitat (Seely Lake/Montana, USA). This mammal moults its coat as colder temperatures, shorter days, and snowfall arrive. In the genetic populations of the temperate forests of the Rocky Mountains and the boreal forests spanning the North American continent, hares that moult from brown to white are abundant (20). However, in coastal areas, and in the third genetic population in the North Pacific, snowfall is brief and less intense, resulting in fewer white individuals. This is due to hybridisation with the black-tailed jackrabbit (Lepus californicus) over 3,000 years ago (17). The hare’s coat has an outer layer, where the longer fur gives each individual its colour, and an inner layer of short fur (19). In winter, the outer layer becomes thicker and denser, while the inner layer maintains a consistent thickness but increases in density. By biomass, the snowshoe hare is the primary herbivore in the North American boreal forest and distinguishes the trophic relationships between continents (21). In Europe, much of the boreal understory remains under snow, providing food for rodents with four-year abundance cycles controlled by small generalist predators (mustelids). In North America, the boreal understory grows above the snow and provides food for hares. In this region, snowshoe hare populations follow 10-year abundance cycles regulated by specialist predators (those that feed almost exclusively on hares), primarily the Canada lynx (Lynx canadensis) (6). Photos courtesy of Alice Kenney and Charles Krebs (Yukon) [see their ecological monitoring program here] and Marketa Zimova (Montana).
However, as the climate changes, springs arrive earlier, winters are delayed, and the frequency and intensity of precipitation have become highly variable. All of this makes it harder for species to adjust their wardrobe to temperature changes (1).
In this context, body colour is a critical factor for birds and mammals that undergo an annual moult (2). In 21 species from the cold latitudes of the Northern Hemisphere, some individuals are brown in summer, but turn white in winter, while others remain brown year round (3). This phenomenon includes weasels, rodents, ptarmigans, foxes, rabbits and hares.
Coral reefs are much more than just a pretty place to visit. They are among the world’s richest ecosystems, hosting about a third of all marine species.
These reefs also directly benefit more than a billion people, providing livelihoods and food security, as well as protection from storms and coastal erosion.
Without coral reefs, the world would be a much poorer place. So when corals die or become damaged, many people try to restore them. But the enormity of the task is growing as the climate keeps warming.
In our new research, we examined the full extent of existing coral restoration projects worldwide. We looked at what drives their success or failure, and how much it would actually cost to restore what’s already been lost. Restoring the reefs we’ve already lost around the world could cost up to A$26 trillion.
Bleached Acropora corals in the Maldives.Davide Seveso/University of Milan
When sea temperatures climb above the seasonal average for sustained periods, corals can become bleached. They lose colour as they expel their symbiotic algae when stressed, revealing the white skeleton underneath. Severe bleaching can kill coral.
Night is the peak activity period for many animal species. In the Western Andes of Ecuador, the Chocó golden scarab flies between forest patches during the night, but urban lighting interferes with their paths and jeopardises populations already struggling to persist in fragmented native forests.
Urban development has created a network of illuminated infrastructure that allows our society to function day and night without interruption. It is no surprise that with so much artificial light, we increasingly have to move farther away from towns and cities to see a sky full of stars.
Light pollution poses a challenge for nocturnal species that have adapted to living in the dimness of night (1, 2) — see documentaries about the impacts of artificial light on wildlife and insects, and a related scientific talk. This problem might be one of the causes of the global decline in insects (3, 4), in turn negatively affecting their role in maintaining agricultural systems through pest control, pollination, and soil quality (5). These concepts are featured by the documentaries The Insect Apocalypse and The Great Death of Insects.
Chocó golden scarab (Chrysina argenteola) walking on forest litter in La Maná (Cotopaxi, Ecuador). Growing to up to 4 cm in length, this species inhabits the tropical rainforest of the Chocó region in the Western Andes (10), where it is frequently attracted to artificial lights at night. The striking colour of this ‘jewel scarab’ is an optical illusion. The exoskeleton is covered with overlapping layers of chitin that polarise light and reflect hues of blue, gold, green, silver, or reddish tones, depending on the species (16). The metallic sheen appears to deter bird predation (17) and might serve as camouflage as well as aid in individual recognition (11). The eyes of insects are ‘compound’ — composed of 100s to 1000s of tubular eyelets (‘ommatidia’), each with its own cornea and lens (18), and all collectively contributing to insect vision. In nocturnal species like the golden scarab, the photoreceptor cells (at the base of each ommatidium) respond more slowly to light compared to diurnal species, allowing the former to collect more nocturnal light per unit of time before forming an image (19). However, just as staring at the sun blinds us, eyes adapted for night vision become overwhelmed by excessive artificial light, disrupting the behaviour of these species. Below the scarab image are two photographs contrasting the day and night landscapes of the same location in Pedro Vicente Maldonado (Pichincha, Ecuador) within the species’ distribution range. Photos courtesy of Martín Bustamante (animal) and Luis Camacho (city).
When flying, nocturnal insects orient their backs toward the sky, using the light of the moon and stars as a reference (6) (explained here and here). However, when they encounter artificial lights, they can no longer distinguish up from down, and so they can become disoriented, flying erratically, like a moth circling a streetlight.
It is estimated that a third of the insects attracted to artificial light die from collisions, burn injuries, exhaustion, and/or predation (7). In the tropics, finding countless dead insects at the base of urban lights is a common scene. Equally important is that artificial light also hinders migration, foraging, and the search for mates in many nocturnal species (1, 8, 9).
Nocturnal jewels
Camacho and collaborators evaluated the effect of artificial lighting at night on the Chocó golden scarab (Chrysina argenteola) (10). This species inhabits the tropical rainforests of the Western Andes from Ecuador to Colombia, and is a member of the group known as ‘jewel scarabs‘ due to their metallic body coloration (11). Because of its nocturnal habits and the larvae’s dependence on wood for food (12), the golden scarab has been increasingly affected by the loss of native forest in combination with light pollution from rural and urban expansion.
Yes, it’s bad, especially for US-based scientists. It also affects scientists in Australia and the rest of the world. But there are ways to get around the problem. There might even be a silver lining to this dark cloud.
Trump cannot stop global climate action, although he might slow it. Nor can he hide the truth by restricting access to data. Climate research will continue despite Trump’s best efforts to hamstring scientists and research institutions.
No strength in ignorance
Last year was the warmest on record, a fact that yet again confirms our worst-case predictions. The world has already surpassed the (arbitrary) 1.5°C threshold increase relative to pre-industrial temperatures — a threshold that only a few years ago we didn’t think we would cross until 2030 at the earliest.
We’re now on track to be living in a world that’s 3°C hotter or more by the end of the century.
But ignoring climate change won’t make it go away. Like the Ministry of Truth in George Orwell’s classic dystopian novel, 1984, Trump seems to believe “ignorance is strength”. He’s trying to erase facts about the climate crisis, perhaps to keep people ignorant and subdued.
What this means for Australian climate science
Many Australian scientists (including me) collaborate regularly with US colleagues, share funding, and publish results together. Knowledge sharing and open-access data are the foundation of advances in science, so Trump’s assault will inevitably slow progress here.
For example, Australian and US scientists regularly collaborate in big-ticket research and policy development related to climate change, such as the Intergovernmental Panel on Climate Change’s Physical Science Basis reports. But even with fewer US scientists in the mix, the research and reporting will continue.
Other reputable climate-data repositories around the world include the European Union’s Climate Data Store, the University of East Anglia’s Climate Research Unit, the Netherlands Meteorological Institute’s Climate Explorer, and the independent WorldClim, to name a few.
While restricting access to US-based websites is inconvenient, we can readily get around the problem. Many of my colleagues have also been downloading data prior to the purge mandate to maintain access.
Consequences for the US
Over the past month I have been inundated with horror stories from many US-based colleagues in academia and the public service, who have lost their jobs and/or research funding. In addition to these very real personal tragedies, the bigger picture is even bleaker.
The loss of scientific and technical expertise these mass sackings entail weakens the capability of the US workforce to discover and develop solutions to climate change. Just when we need good scientific and engineering innovations more than ever, a massive capacity is being erased before our eyes.
More emissions mean more climate change, especially when you’re already one of the biggest contributors to the global problem. The US is the second-highest greenhouse emitter in the world, behind only China.
On his first day as president, Trump withdrew the US from the Paris climate agreement. This effectively removes his country from all binding limits on actions that contribute to climate change.
Weakening international treaties is a two-edged sword, because it not only lets the US off the leash, it also potentially discourages other nations from acting responsibly. Analogous to the “unresponsive bystander effect”, many nations may now be more hesitant to commit to reductions because one of the biggest emitters refuses to do anything about it.
Trump has also slashed US international aid, which will slow climate action in countries that need the most assistance.
Overall, faster rates of warming will inevitably put more strain on natural resources and agricultural production. This could increase the probability of international warfare over water, food and other essential natural resources. Because autocratic countries cope worse with food shortages than democratic ones, climate emergencies will penalise nations led by despots more heavily.
Trump’s foolhardy anti-climate campaign is enough to make many people despair. But there are a few faint glimmers of hope on the horizon.
As the US shirks its domestic and international responsibilities, other countries might resolve to do more. Not relying on the US could force capacity-building elsewhere. Some even suggest without the US at the table slowing progress, stronger climate action might result.
Americans have their own daunting fight on their hands. But the rest of the world will have to take up the slack if we have any chance of limiting the health, wealth, equality, human rights and biodiversity calamities now unfolding because of climate change.
Corey J. A. Bradshaw, Matthew Flinders Professor of Global Ecology and Node Leader in the ARC Centre of Excellence for Indigenous and Environmental Histories and Futures, Flinders University
If you’re like me, you use a lot of loops in R. I do not profess to be the most efficient coder, but loops make sense to me and I’m generally not concerned about make the fastest simulations.
But sometimes my loops take some time to finish, so I often add a rolling text update during the simulation to know how far it has progressed. But of course, I have to look at the R console to see how far things have come. Being a bit away from the central tendency of the spectrum, I can get absorbed in doing other things, so I often miss when the simulation is complete.
In a fit of excess geekiness, I’ve recently discovered voice prompts in MacOS that I can now code directly into my R simulations to give verbal updates on their progress. I find these immensely useful. I’ve therefore decided to share the basic code, because I know some other geeks out there might also appreciate the tool. Apologies — I haven’t investigated how to do this in a PC environment, so the following examples are MacOS-specific.
First, go to your Accessibility settings in System Settings in your Mac. Click on System Voice to see what voices you have access to, and which voices you wish to download to your machine. There are many languages supported.
When you construct a loop in R, add the following code within and before the loop content (this example is in English):
iter <- 1000 # number of iterations
itdiv <- iter/100 # iteration divisor 1
itdiv2 <- iter/10 # iteration divisor 2
st.time <- Sys.time() # time at start of simulation
# loop from 1 to iter
for (i in 1:iter) {
# pause execution for 0.05 seconds (this would normally be the guts of your loop functions)
Sys.sleep(0.05)
# loop updaters with voice (English)
if (i %% itdiv==0) print(paste("iter = ", i, sep=""))
if (i %% itdiv2==0 & i < iter) system2("say", c("-v", "Fiona", paste(round(100*(i/iter), 0),
"per cent complete"))) # updates every 10% complete
if (i == 0.95*iter) system2("say", c("-v", "Fiona", paste(round(100*(i/iter), 0),
"per cent complete"))) # announce at 95% complete
if (i == 0.99*iter) system2("say", c("-v", "Fiona", paste(round(100*(i/iter), 0),
"per cent complete"))) # announce at 99% complete
if (i == iter) system2("say", c("-v", "Lee", "simulation complete"))
if (i == iter) system2("say", c("-v", "Lee", paste(round(as.numeric(Sys.time() - st.time,
units = "mins"), 2), "minutes elapsed")))
}
Here I’ve used the female Scottish voice ‘Fiona’ and the male Australian voice ‘Lee’.
Quite a bit late this year, but I’ve finally put together the 2023 conservation / ecology / sustainability journal ranks based on my (published) journal-ranking method (as I’ve done every year since 2008).
After 16 years of doing this exercise, I can’t help but notice that most journals don’t do much differently from year to year. They mostly tend to publish the same number of papers, get the same number of total publications, and therefore, remain approximately in the same rank relative to others.
Some things to note: Clarivate continues to modify its algorithm, meaning that most journal Impact Factors have gone down yet again. This is somewhat irrelevant from the perspective of relative ranking, but it might piss off a few journals.
I therefore present the new 2023 ranks for: (i) 111 ecology, conservation and multidisciplinary journals, (ii) 29 open-access (i.e., you have to pay) journals from the previous category, (iii) 68 ‘ecology’ journals, (iv) 33 ‘conservation’ journals, (v) 44 ‘sustainability’ journals (with general and energy-focussed journals included), and (vi) 21 ‘marine & freshwater’ journals.
Human overpopulation is often depicted in the media in one of two ways: as either a catastrophic disaster or an overly-exaggerated concern. Yet the data understood by scientists and researchers is clear. So what is the actual state of our overshoot, and, despite our growing numbers, are we already seeing the signs that the sixth mass extinction is underway?
In a recent episode of The Great Simplification podcast, Nate Hagens was joined by global ecologist Corey Bradshaw to discuss his recent research on the rapid decline in biodiversity, how population and demographics will change in the coming decades, and what both of these will mean for complex global economies currently reliant on a stable environment.
Australia is home to about one in 12 of the world’s species of animals, birds, plants and insects – between 600,000 and 700,000 species. More than 80% of Australian plants and mammals and just under 50% of our birds are found nowhere else.
But habitat destruction, climate change, and invasive species are wreaking havoc on Earth’s rich biodiversity, and Australia is no exception.
More and more species stand on the edge of oblivion. That’s just the ones we know enough about to list formally as threatened. Many more are in trouble, especially in the oceans. Change is the new constant. As the world heats up and ecosystems warp, new combinations of species can emerge without an evolutionary connection, creating novel communities.
It is still possible to stop species from dying out. But it will take an unprecedented effort.
The vulnerable southern bell (growling grass) frog (Litoria raniformis). Rupert Mathwin/Flinders University
If several fossils of an extinct population or species are dated, we can estimate how long ago the extinction event took place. In our new paper, we describe CRIWM, a new method to estimate extinction time using times series of fossils whose ages have been measured by radiocarbon dating.And yes, there’s an R package — Rextinct — to go with that!
While the Earth seems to gather all the conditions for life to thrive, over 99.9% of all species that ever lived are extinct today. From a distance, pristine landscapes might look similar today and millennia ago: blue seas with rocky and sandy coasts and grasslands and mountain ranges watered by rivers and lakes and covered in grass, bush and trees.
But zooming in, the picture is quite different because species identities have never stopped changing — with ‘old’ species being slowly replaced by ‘new’ ones. Fortunately, much like the collection of books in the library summarises the history of literature, the fossilised remnants of extinct organisms represent an archive of the kinds of creatures that have ever lived. This fossil record can be used to determine when and why species disappear. In that context, measuring the age of fossils is a useful task for studying the history of biodiversity and its connections to the planet’s present.
In our new paper published in the journal Quaternary Geochronology (1), we describe CRIWM (calibration-resampled inverse-weighted McInerny), a statistical method to estimate extinction time using times series of fossils that have been dated using radiocarbon dating.
Why radiocarbon dating? Easy. It is the most accurate and precise chronometric method to date fossils younger than 50,000 to 55,000 years old (2, 3). This period covers the Holocene (last 11,700 years or so), and the last stretch of the late Pleistocene (~ 130,000 years ago to the Holocene), a crucial window of time witnessing the demise of Quaternary megafauna at a planetary scale (4) (see videos here, here and here), and the global spread of anatomically modern humans (us) ‘out of Africa’ (see here and here).
Why do we need a statistical method? Fossilisation (the process of body remains being preserved in the rock record) is rare and finding a fossil is so improbable that we need maths to control for the incompleteness of the fossil record and how this fossil record relates to the period of survival of an extinct species.
A brief introduction to radiocarbon dating
First, let’s revise the basics of radiocarbon dating (also explained here and here). This chronometric technique measures the age of carbon-rich organic materials — from shells and bones to the plant and animal components used to write an ancient Koran, make a wine vintage and paint La Mona Lisa and Neanderthal caves.
A global database set up by scientists to assemble data on the economic cost of biological invasions in support of effective government management strategies has grown to include all known invasive species.
Now involving 145 researchers from 44 countries — the current version of InvaCost has 13,553 entries in 22 languages and enables scientists to develop a clear picture about the major threats globally of invasive species to ecosystems, biodiversity, and human well-being.
Biological invasions are caused by species introduced on purpose or accidentally by humans to areas outside of their natural ranges. From cats and weeds, to crop pests and diseases, invasive species are a worldwide scourge.
Invasive species have cost over US$2 trillion globally since the 1970s by damaging goods and services, and through the costs of managing them, and these economic costs are only increasing.
A new synthesis published in the journal BioSciencedocuments the progress of the InvaCost endeavour.The study provides a timeline of the state of invasion costs, starting with prior flaws and shortcomings in the scientific literature, then how InvaCost has helped to alleviate and address these issues, and what the future potentially holds for research and policymakers.
In light of new genetic research on the identity of ‘wild dogs’ and dingoes across Australia, the undersigned wish to express concern with current South Australia Government policy regarding the management and conservation of dingoes. Advanced DNA research on dingoes has demonstrated that dingo-dog hybridisation is much less common than thought, that most DNA tested dingoes had little domestic dog ancestry and that previous DNA testing incorrectly identified many dingoes as hybrids (Cairns et al. 2023). We have serious concerns about the threat current South Australian public policy poses to the survival of the ‘Big Desert’ dingo population found in Ngarkat Conservation Park and surrounding areas.
We urge the South Australian Government to:
Revoke the requirement that all landholders follow minimum baiting standards, including organic producers or those not experiencing stock predation. Specifically
Dingoes in Ngarkat Conservation park (Region 4) should not be destroyed or subjected to ground baiting and trapping every 3 months. The Ngarkat dingo population is a unique and isolated lineage of dingo that is threatened by inbreeding and low genetic diversity. Dingoes are a native species and all native species should be protected inside national parks and conservation areas.
Landholders should not be required to carry out ground baiting on land if there is no livestock predation occurring. Furthermore, landholders should be supported to adopt non-lethal tools and strategies to mitigate the risk of livestock predation including the use of livestock guardian animals, which are generally incompatible with ground and aerial 1080 baiting.
Revoke permission for aerial baiting of dingoes (incorrectly called “wild dogs”) in all Natural Resource Management regions – including within national parks. Native animals should be protected in national parks and conservation areas.
Cease the use of inappropriate and misleading language to label dingoes as “wild dogs”. Continued use of the term “wild dogs” is not culturally respectful to First Nations peoples and is not evidence-based.
Proactively engage with First Nations peoples regarding the management of culturally significant species like dingoes. For example, the Wotjobaluk nation should be included in consultation regarding the management of dingoes in Ngarkat Conservation Park.
Changes in South Australia public policy are justified based on genetic research by Cairns et al. (2023) that overturns previous misconceptions about the genetic status of dingoes. It demonstrates:
Most “wild dogs” DNA tested in arid and remote parts of Australia were dingoes with no evidence of dog ancestry. There is strong evidence that dingo-dog hybridisation is uncommon, with firstcross dingo-dog hybrids and feral dogs rarely being observed in the wild. In Ngarkat Conservation park none of DNA tested animals had evidence of domestic dog ancestry, all were ‘pure’ dingoes.
Previous DNA testing methods misidentified pure dingoes as being mixed. All previous genetic surveys of wild dingo populations used a limited 23-marker DNA test. This is the method currently used by NSW Department of Primary Industries, which DNA tests samples from NSW Local Land Services, National Parks and Wildlife Service, and other state government agencies. Comparisons of DNA testing methods find that the 23-marker DNA test frequently misidentified animals as dingo-dog hybrids. Existing knowledge of dingo ancestry across South Australia, particularly from Ngarkat Conservation park is incorrect; policy needs to be based on updated genetic surveys.
There are multiple dingo populations in Australia. High-density genomic data identified more than four wild dingo populations in Australia. In South Australia there are at least two dingo populations present: West and Big Desert. The West dingo population was observed in northern South Australia, but also extends south of the dingo fence. The Big Desert population extends from Ngarkat Conservation park in South Australia into the Big Desert and Wyperfield region of Victoria.
The Ngarkat Dingo population is threatened by low genetic variability. Preliminary evidence from high density genomic testing of dingoes in Ngarkat Conservation park and extending into western Victoria found evidence of limited genetic variability which is a serious conservation concern. Dingoes in Ngarkat and western Victoria had extremely low genetic variability and no evidence of gene flow with other dingo populations, demonstrating their effective isolation. This evidence suggests that the Ngarkat (and western Victorian) dingo population is threatened by inbreeding and genetic isolation. Continued culling of the Ngarkat dingo population will exacerbate the low genetic variability and threatens the persistence of this population.
But just how far-fetched is it? Could pathogens that were once common on Earth – but frozen for millennia in glaciers, ice caps and permafrost – emerge from the melting ice to lay waste to modern ecosystems? The potential is, in fact, quite real.
In 2014, a giant “zombie” Pithovirus sibericum virus was revived from 30,000-year-old Siberian permafrost.
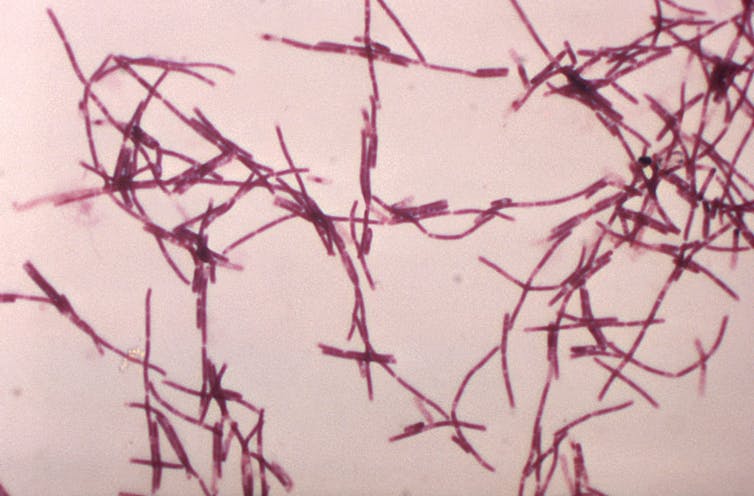
And in 2016, an outbreak of anthrax (a disease caused by the bacterium Bacillus anthracis) in western Siberia was attributed to the rapid thawing of B. anthracis spores in permafrost. It killed thousands of reindeer and affected dozens of people.
Bacillus anthracis is a soil bacterium that causes anthrax. William A. Clark/USCDCP
More recently, scientists found remarkable genetic compatibility between viruses isolated from lake sediments in the high Arctic and potential living hosts.
Earth’s climate is warming at a spectacular rate, and up to four times faster in colder regions such as the Arctic. Estimates suggest we can expect four sextillion (4,000,000,000,000,000,000,000) microorganisms to be released from ice melt each year. This is about the same as the estimated number of stars in the universe.
However, despite the unfathomably large number of microorganisms being released from melting ice (including pathogens that can potentially infect modern species), no one has been able to estimate the risk this poses to modern ecosystems.
In a new study published today in the journal PLOS Computational Biology, we calculated the ecological risks posed by the release of unpredictable ancient viruses.
As I’ve done every year for the last 15 years, I can now present the 2022 conservation / ecology / sustainability journal ranks based on my (published) journal-ranking method.
Although both the Clarivate (Impact Factor, Journal Citation Indicator, Immediacy Index) and Scopus (CiteScore, Source-Normalised Impact Per Paper, SCImago Journal Rank) values have been out for about a month or so, the Google (h5-index, h5-median) scores only came out yesterday.
This year’s also a bit weird from the perspective of the Clarivate ranks. First, Impact Factors will no longer be provided to three significant digits, but only to one (e.g., 7.2 versus 7.162). That’s not such a big deal, but it does correct for relative ranks based on false precision. However, the biggest changes are more methdological — Impact Factors now take online articles into account (in the denominator), so most journals will have a lower Impact Factor this year compared to last. In fact, of the 105 journals in the ecology/conservation/multidisciplinary category that have data for both 2021 and 2022, the 2022 Impact Factors are a median 15% lower than the 2021 values.
Another effect in play appears to have been the pandemic. The worst of the pandemic happened right during the assessment period, and I’m pretty sure this is reflected both in terms of the number of articles published (down a median of 10%) and total number of citations in the assessment period (down 7%) per journal.
But using my method, these changes a somewhat irrelevant because I calculate relative ranks, not an absolute score.
I therefore present the new 2022 ranks for: (i) 108 ecology, conservation and multidisciplinary journals, (ii) 28 open-access (i.e., you have to pay) journals from the previous category, (iii) 66 ‘ecology’ journals, (iv) 31 ‘conservation’ journals, (v) 43 ‘sustainability’ journals (with general and energy-focussed journals included), and (vi) 21 ‘marine & freshwater’ journals.
Have you ever watched a nature documentary and marvelled at the intricate dance of life unfolding on screen? From the smallest insect to the largest predator, every creature plays a role in the grand performance of our planet’s biosphere. But what happens when one of these performers disappears?
In this post, we delve into our recent article Estimating co-extinction risks in terrestrial ecosystems just published in Global Change Biology, in which we discuss the cascading effects of species loss and the risks of ‘co-extinction’.
But what does ‘co-extinction’ really mean?
Imagine an ecosystem as a giant web of interconnected species. Each thread represents a relationship between two species — for example, a bird that eats a certain type of insect, or a plant that relies on a specific species of bee for pollination. Now, what happens if one of these species in the pair disappears? The thread breaks and the remaining species loses an interaction. This could potentially lead to its co-extinction, which is essentially the domino effect of multiple species losses in an ecosystem.
A famous example of this effect can be seen with the invasion of the cane toad (Rhinella marina) across mainland Australia, which have caused trophic cascades and species compositional changes in these communities.
The direct extinction of one species, caused by effects such as global warming for example, has the potential to cause other species also to become extinct indirectly.
We’ve just published a new paper showing that young red kangaroos (Osphranter rufus) protected by the dingo-proof fence take more time to grow up than their counterparts on the other side, who quickly outgrow the risk of being a dingo’s next meal.
Published in the Journal of Mammalogy, our article led by Rex Mitchell also revealed that there are more young and female kangaroos inside the dingo-proof fence, showing that the fence impacts on different aspects of the red kangaroo’s life cycle.
Red kangaroos are one of the dingo’s favourite prey species, so it’s not surprising to find fewer of the smaller females and younger animals when there are more dingoes around. However, we didn’t expect that young animals inside the fence were lighter and smaller than those outside the fence. Read the rest of this entry »
Those of us living with cats share our homes with an ancestral predator, one adapted for hunting and the frequent, exclusive consumption of meat. These instincts become fully activated outside the domestic environment, where cats pose a global threat to wildlife. Pets are family. We celebrate their arrival with the same joy as a grand…
Many animals avoid contact with people. In protected areas of the African savanna, mammals flee more intensely upon hearing human conversations than when they hear lions or sounds associated with hunting. This fear of humans affects how species use and move in their habitat. Throughout our lives, we interact with hundreds of wildlife species without…
Deep-sea sharks include some of the longest-lived vertebrates known. The record holder is the Greenland shark, with a recently estimated maximum age of nearly 400 years. Their slow life cycle makes them vulnerable to fisheries. Humans rarely live longer than 100 years. But many other animals and plants can live for several centuries or even millennia, particularly…