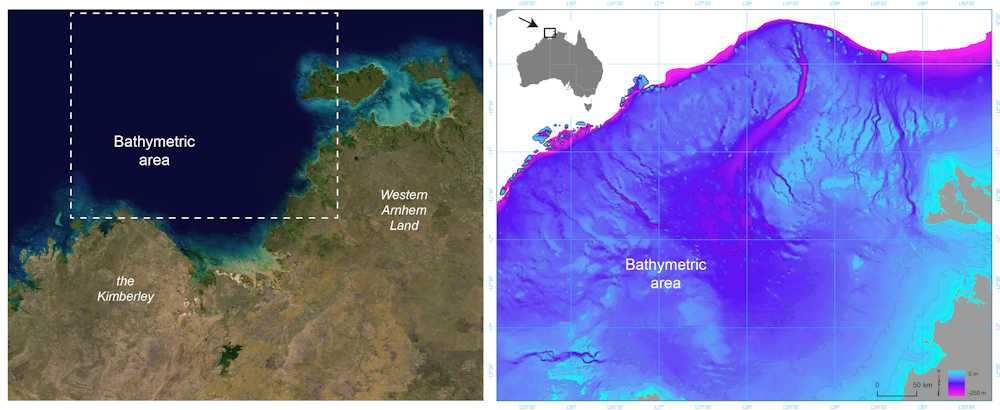
For much of the 65,000 years of Australia’s human history, the now-submerged northwest continental shelf connected the Kimberley and western Arnhem Land. This vast, habitable realm covered nearly 390,000 square kilometres, an area one-and-a-half times larger than New Zealand is today.
Left: Satellite image of the submerged northwest shelf region. Right: Drowned landscape map of the study area. US Geological Survey, Geoscience Australia
It was likely a single cultural zone, with similarities in ground stone-axe technology, styles of rock art, and languages found by archaeologists in the Kimberley and Arnhem Land.
There is plenty of archaeological evidence humans once lived on continental shelves – areas that are now submerged – all around the world. Such hard evidence has been retrieved from underwater sites in the North Sea, Baltic Sea and Mediterranean Sea, and along the coasts of North and South America, South Africa and Australia.
In a newly published study in Quaternary Science Reviews, we reveal details of the complex landscape that existed on the Northwest Shelf of Australia. It was unlike any landscape found on our continent today.
A continental split
Around 18,000 years ago, the last ice age ended. Subsequent warming caused sea levels to rise and drown huge areas of the world’s continents. This process split the supercontinent of Sahul into New Guinea and Australia, and cut Tasmania off from the mainland.
Unlike in the rest of the world, the now-drowned continental shelves of Australia were thought to be environmentally unproductive and little used by First Nations peoples.
But mounting archaeological evidence shows this assumption is incorrect. Many large islands off Australia’s coast – islands that once formed part of the continental shelves – show signs of occupation before sea levels rose.
Stone tools have also recently been found on the sea floor off the coast of the Pilbara region of Western Australia.
If several fossils of an extinct population or species are dated, we can estimate how long ago the extinction event took place. In our new paper, we describe CRIWM, a new method to estimate extinction time using times series of fossils whose ages have been measured by radiocarbon dating.And yes, there’s an R package — Rextinct — to go with that!
While the Earth seems to gather all the conditions for life to thrive, over 99.9% of all species that ever lived are extinct today. From a distance, pristine landscapes might look similar today and millennia ago: blue seas with rocky and sandy coasts and grasslands and mountain ranges watered by rivers and lakes and covered in grass, bush and trees.
But zooming in, the picture is quite different because species identities have never stopped changing — with ‘old’ species being slowly replaced by ‘new’ ones. Fortunately, much like the collection of books in the library summarises the history of literature, the fossilised remnants of extinct organisms represent an archive of the kinds of creatures that have ever lived. This fossil record can be used to determine when and why species disappear. In that context, measuring the age of fossils is a useful task for studying the history of biodiversity and its connections to the planet’s present.
In our new paper published in the journal Quaternary Geochronology (1), we describe CRIWM (calibration-resampled inverse-weighted McInerny), a statistical method to estimate extinction time using times series of fossils that have been dated using radiocarbon dating.
Why radiocarbon dating? Easy. It is the most accurate and precise chronometric method to date fossils younger than 50,000 to 55,000 years old (2, 3). This period covers the Holocene (last 11,700 years or so), and the last stretch of the late Pleistocene (~ 130,000 years ago to the Holocene), a crucial window of time witnessing the demise of Quaternary megafauna at a planetary scale (4) (see videos here, here and here), and the global spread of anatomically modern humans (us) ‘out of Africa’ (see here and here).
Why do we need a statistical method? Fossilisation (the process of body remains being preserved in the rock record) is rare and finding a fossil is so improbable that we need maths to control for the incompleteness of the fossil record and how this fossil record relates to the period of survival of an extinct species.
A brief introduction to radiocarbon dating
First, let’s revise the basics of radiocarbon dating (also explained here and here). This chronometric technique measures the age of carbon-rich organic materials — from shells and bones to the plant and animal components used to write an ancient Koran, make a wine vintage and paint La Mona Lisa and Neanderthal caves.
A global database set up by scientists to assemble data on the economic cost of biological invasions in support of effective government management strategies has grown to include all known invasive species.
Now involving 145 researchers from 44 countries — the current version of InvaCost has 13,553 entries in 22 languages and enables scientists to develop a clear picture about the major threats globally of invasive species to ecosystems, biodiversity, and human well-being.
Biological invasions are caused by species introduced on purpose or accidentally by humans to areas outside of their natural ranges. From cats and weeds, to crop pests and diseases, invasive species are a worldwide scourge.
Invasive species have cost over US$2 trillion globally since the 1970s by damaging goods and services, and through the costs of managing them, and these economic costs are only increasing.
A new synthesis published in the journal BioSciencedocuments the progress of the InvaCost endeavour.The study provides a timeline of the state of invasion costs, starting with prior flaws and shortcomings in the scientific literature, then how InvaCost has helped to alleviate and address these issues, and what the future potentially holds for research and policymakers.
In light of new genetic research on the identity of ‘wild dogs’ and dingoes across Australia, the undersigned wish to express concern with current South Australia Government policy regarding the management and conservation of dingoes. Advanced DNA research on dingoes has demonstrated that dingo-dog hybridisation is much less common than thought, that most DNA tested dingoes had little domestic dog ancestry and that previous DNA testing incorrectly identified many dingoes as hybrids (Cairns et al. 2023). We have serious concerns about the threat current South Australian public policy poses to the survival of the ‘Big Desert’ dingo population found in Ngarkat Conservation Park and surrounding areas.
We urge the South Australian Government to:
Revoke the requirement that all landholders follow minimum baiting standards, including organic producers or those not experiencing stock predation. Specifically
Dingoes in Ngarkat Conservation park (Region 4) should not be destroyed or subjected to ground baiting and trapping every 3 months. The Ngarkat dingo population is a unique and isolated lineage of dingo that is threatened by inbreeding and low genetic diversity. Dingoes are a native species and all native species should be protected inside national parks and conservation areas.
Landholders should not be required to carry out ground baiting on land if there is no livestock predation occurring. Furthermore, landholders should be supported to adopt non-lethal tools and strategies to mitigate the risk of livestock predation including the use of livestock guardian animals, which are generally incompatible with ground and aerial 1080 baiting.
Revoke permission for aerial baiting of dingoes (incorrectly called “wild dogs”) in all Natural Resource Management regions – including within national parks. Native animals should be protected in national parks and conservation areas.
Cease the use of inappropriate and misleading language to label dingoes as “wild dogs”. Continued use of the term “wild dogs” is not culturally respectful to First Nations peoples and is not evidence-based.
Proactively engage with First Nations peoples regarding the management of culturally significant species like dingoes. For example, the Wotjobaluk nation should be included in consultation regarding the management of dingoes in Ngarkat Conservation Park.
Changes in South Australia public policy are justified based on genetic research by Cairns et al. (2023) that overturns previous misconceptions about the genetic status of dingoes. It demonstrates:
Most “wild dogs” DNA tested in arid and remote parts of Australia were dingoes with no evidence of dog ancestry. There is strong evidence that dingo-dog hybridisation is uncommon, with firstcross dingo-dog hybrids and feral dogs rarely being observed in the wild. In Ngarkat Conservation park none of DNA tested animals had evidence of domestic dog ancestry, all were ‘pure’ dingoes.
Previous DNA testing methods misidentified pure dingoes as being mixed. All previous genetic surveys of wild dingo populations used a limited 23-marker DNA test. This is the method currently used by NSW Department of Primary Industries, which DNA tests samples from NSW Local Land Services, National Parks and Wildlife Service, and other state government agencies. Comparisons of DNA testing methods find that the 23-marker DNA test frequently misidentified animals as dingo-dog hybrids. Existing knowledge of dingo ancestry across South Australia, particularly from Ngarkat Conservation park is incorrect; policy needs to be based on updated genetic surveys.
There are multiple dingo populations in Australia. High-density genomic data identified more than four wild dingo populations in Australia. In South Australia there are at least two dingo populations present: West and Big Desert. The West dingo population was observed in northern South Australia, but also extends south of the dingo fence. The Big Desert population extends from Ngarkat Conservation park in South Australia into the Big Desert and Wyperfield region of Victoria.
The Ngarkat Dingo population is threatened by low genetic variability. Preliminary evidence from high density genomic testing of dingoes in Ngarkat Conservation park and extending into western Victoria found evidence of limited genetic variability which is a serious conservation concern. Dingoes in Ngarkat and western Victoria had extremely low genetic variability and no evidence of gene flow with other dingo populations, demonstrating their effective isolation. This evidence suggests that the Ngarkat (and western Victorian) dingo population is threatened by inbreeding and genetic isolation. Continued culling of the Ngarkat dingo population will exacerbate the low genetic variability and threatens the persistence of this population.
But just how far-fetched is it? Could pathogens that were once common on Earth – but frozen for millennia in glaciers, ice caps and permafrost – emerge from the melting ice to lay waste to modern ecosystems? The potential is, in fact, quite real.
In 2014, a giant “zombie” Pithovirus sibericum virus was revived from 30,000-year-old Siberian permafrost.
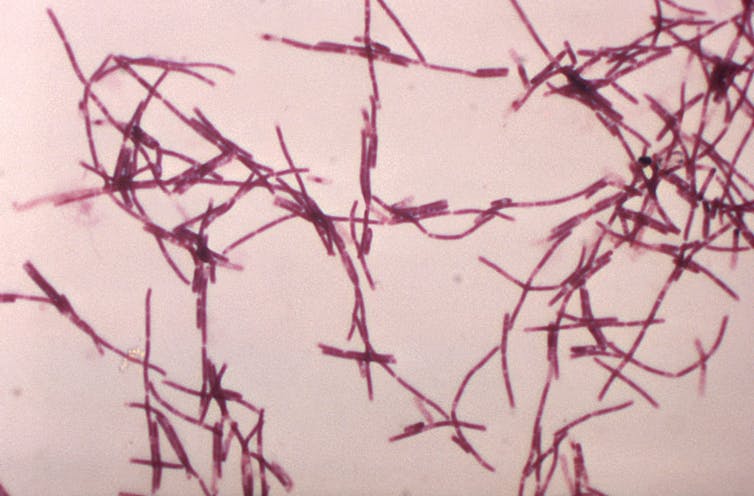
And in 2016, an outbreak of anthrax (a disease caused by the bacterium Bacillus anthracis) in western Siberia was attributed to the rapid thawing of B. anthracis spores in permafrost. It killed thousands of reindeer and affected dozens of people.
Bacillus anthracis is a soil bacterium that causes anthrax. William A. Clark/USCDCP
More recently, scientists found remarkable genetic compatibility between viruses isolated from lake sediments in the high Arctic and potential living hosts.
Earth’s climate is warming at a spectacular rate, and up to four times faster in colder regions such as the Arctic. Estimates suggest we can expect four sextillion (4,000,000,000,000,000,000,000) microorganisms to be released from ice melt each year. This is about the same as the estimated number of stars in the universe.
However, despite the unfathomably large number of microorganisms being released from melting ice (including pathogens that can potentially infect modern species), no one has been able to estimate the risk this poses to modern ecosystems.
In a new study published today in the journal PLOS Computational Biology, we calculated the ecological risks posed by the release of unpredictable ancient viruses.
As I’ve done every year for the last 15 years, I can now present the 2022 conservation / ecology / sustainability journal ranks based on my (published) journal-ranking method.
Although both the Clarivate (Impact Factor, Journal Citation Indicator, Immediacy Index) and Scopus (CiteScore, Source-Normalised Impact Per Paper, SCImago Journal Rank) values have been out for about a month or so, the Google (h5-index, h5-median) scores only came out yesterday.
This year’s also a bit weird from the perspective of the Clarivate ranks. First, Impact Factors will no longer be provided to three significant digits, but only to one (e.g., 7.2 versus 7.162). That’s not such a big deal, but it does correct for relative ranks based on false precision. However, the biggest changes are more methdological — Impact Factors now take online articles into account (in the denominator), so most journals will have a lower Impact Factor this year compared to last. In fact, of the 105 journals in the ecology/conservation/multidisciplinary category that have data for both 2021 and 2022, the 2022 Impact Factors are a median 15% lower than the 2021 values.
Another effect in play appears to have been the pandemic. The worst of the pandemic happened right during the assessment period, and I’m pretty sure this is reflected both in terms of the number of articles published (down a median of 10%) and total number of citations in the assessment period (down 7%) per journal.
But using my method, these changes a somewhat irrelevant because I calculate relative ranks, not an absolute score.
I therefore present the new 2022 ranks for: (i) 108 ecology, conservation and multidisciplinary journals, (ii) 28 open-access (i.e., you have to pay) journals from the previous category, (iii) 66 ‘ecology’ journals, (iv) 31 ‘conservation’ journals, (v) 43 ‘sustainability’ journals (with general and energy-focussed journals included), and (vi) 21 ‘marine & freshwater’ journals.
Have you ever watched a nature documentary and marvelled at the intricate dance of life unfolding on screen? From the smallest insect to the largest predator, every creature plays a role in the grand performance of our planet’s biosphere. But what happens when one of these performers disappears?
In this post, we delve into our recent article Estimating co-extinction risks in terrestrial ecosystems just published in Global Change Biology, in which we discuss the cascading effects of species loss and the risks of ‘co-extinction’.
But what does ‘co-extinction’ really mean?
Imagine an ecosystem as a giant web of interconnected species. Each thread represents a relationship between two species — for example, a bird that eats a certain type of insect, or a plant that relies on a specific species of bee for pollination. Now, what happens if one of these species in the pair disappears? The thread breaks and the remaining species loses an interaction. This could potentially lead to its co-extinction, which is essentially the domino effect of multiple species losses in an ecosystem.
A famous example of this effect can be seen with the invasion of the cane toad (Rhinella marina) across mainland Australia, which have caused trophic cascades and species compositional changes in these communities.
The direct extinction of one species, caused by effects such as global warming for example, has the potential to cause other species also to become extinct indirectly.
We’ve just published a new paper showing that young red kangaroos (Osphranter rufus) protected by the dingo-proof fence take more time to grow up than their counterparts on the other side, who quickly outgrow the risk of being a dingo’s next meal.
Published in the Journal of Mammalogy, our article led by Rex Mitchell also revealed that there are more young and female kangaroos inside the dingo-proof fence, showing that the fence impacts on different aspects of the red kangaroo’s life cycle.
Red kangaroos are one of the dingo’s favourite prey species, so it’s not surprising to find fewer of the smaller females and younger animals when there are more dingoes around. However, we didn’t expect that young animals inside the fence were lighter and smaller than those outside the fence. Read the rest of this entry »
The way that eels migrate along rivers and seas is mesmerising. There has been scientific agreement since the turn of the 20th Century that the Sargasso Sea is the breeding home to the sole European species. But it has taken more than two centuries since Carl Linnaeus gave this snake-shaped fish its scientific name before an adult was discovered in the area where they mate and spawn.
Even among nomadic people, the average human walks no more than a few dozen kilometres in a single trip. In comparison, the animal kingdom is rife with migratory species that traverse continents, oceans, and even the entire planet (1).
The European eel (Anguilla anguilla) is an outstanding example. Adults migrate up to 5000 km from the rivers and coastal wetlands of Europe and northern Africa to reproduce, lay their eggs, and die in the Sargasso Sea — an algae-covered sea delimited by oceanic currents in the North Atlantic.
The European eel (Anguilla Anguilla) is an omnivorous fish that migrates from European and North African rivers to the Sargasso Sea to mate and die (18). Each individual experiences 4 distinct developmental phases, which look so different that they have been described as three distinct species (19): A planktonic, leaf-like larva (ilecocephalus phase) emerges from each egg and takes up to 3 years to cross the Atlantic. Off the Afro-European coasts, the larva transforms into a semi-transparent tiny eel (iiglass phase) that enters wetlands and estuaries, and travels up the rivers as it gains weight and pigment (iiiyellow phase). They remain there for up to 20 years, rarely growing larger than 1 m in length and 4 kg in weight (females are larger than males) — see underwater footage here and here. Sexual maturity ultimately begins to adjust to the migration to the sea: a darker, saltier, and deeper environment than the river. Their back and belly turn bronze and silver (ivsilver phase), respectively, the eyes increase in size and the number of photoreceptors multiplies (function = submarine vision), the stomach shrinks and loses its digestive function, the walls of the swim bladder thicken (function = floating in the water column), and the fat content of tissues increases by up to 30% of body weight (function = fuel for transoceanic travelling). And finally, the reproductive system will gradually develop while eels navigate to the Sargasso Sea — a trip during which they fast. Photos courtesy of Sune Riis Sørensen (2-day embryo raised at www.eel-hatch.dk and leptocephalus from the Sargasso Sea) and Lluís Zamora (Ter River, Girona, Spain: glass eels in Torroella de Montgrí, 70 cm yellow female in Bonmatí, and 40 cm silver male showing eye enlargement in Bescanó). Eggs and sperm are only known from in vitro fertilisation in laboratories and fish farms (20).
As larvae emerge, they drift with the prevailing marine currents over the Atlantic to the European and African coasts (2). The location of the breeding area was unveiled in the early 20th Century as a result of the observation that the size of the larvae caught in research surveys gradually decreased from Afro-European land towards the Sargasso Sea (3, 4). Adult eels had been tracked by telemetry in their migration route converging on the Azores Archipelago (5), but none had been recorded beyond until recently.
Crossing the Atlantic
To complete this piece of the puzzle, Rosalind Wright and collaborators placed transmitters in 21 silver females and released them in the Azores (6). These individuals travelled between 300 and 2300 km, averaging 7 km each day. Five arrived in the Sargasso Sea, and one of them, after a swim of 243 days (from November 2019 to July 2020), reached what for many years had been the hypothetical core of the breeding area (3, 4). It is the first direct record of a European eel ending its reproductive journey.
Eels use the magnetic fields in their way back to the Sargasso Sea and rely on an internal compass that records the route they made as larvae (7). The speed of navigation recorded by Wright is slower than in many long-distance migratory vertebrates like birds, yet it is consistent across the 16 known eel species (8).
Telemetry (6) and fisheries (14) of European eel (Anguilla anguilla). Eel silhouettes indicate the release point of 21 silver females in Azores in 2018 (orange) and 2019 (yellow), the circles show the position where their transmitters stopped sending signals, and the grey background darkens with water depth. The diagrams display the distance travelled and the speed per eel, where the circle with bold border represents the female that reached the centre of the hypothetical spawning area in the Sargasso Sea (dashed lines in the map) (3). Blue, green and pink symbols indicate the final location of eels equipped with teletransmitters in previous studies, finding no individual giving location signals beyond the Azores Archipelago (6). The barplot shows commercial catches (1978-2021) of yellow+silver eels in those European countries with historical landings exceeding 30,000 t (no data available for France prior to 1986), plus Spain (6120 t from 1951) — excluding recreational fishery and farming which, in 2020, totalled 300 and 4600 t, respectively (14). Red circles represent glass-eel catches added up for France (> 90% of all-country landings), Great Britain, Portugal, and Spain. Catches have kept declining since the 1980s. One kg of glass eels contains some 3000 individuals, so the glass-eel fishery has a far greater impact on stocks than the adult fishery.
Wright claimed that, instead of swiftly migrating for early spawning, eels engage in a protracted migration at depth. This behaviour serves to conserve their energy and minimises the risk of dying (6). The delay also allows them to reach full reproductive potential since, during migration, eels stop eating and mobilise all their resources to swim and reproduce (9).
Other studies have revealed that adults move in deep waters in daylight but in shallow waters at night, and that some individuals are faster than others (3 to 47 km per day) (5). Considering that (i) this fish departs Europe and Africa between August and December and (ii) spawning occurs in the Sargasso Sea from December to May, it is unknown whether different individuals might breed 1 or 2 years after they begin their oceanic migration.
Management as complex as life itself
The European eel started showing the first signs of decline at the end of the 19th Century (10, 11). In 2008, the species was listed as Critically Endangered by the IUCN, and its conservation status has since remained in that category — worse than that of the giant panda (Ailuropoda melanoleuca) or the Iberian lynx (Lynx pardinus).
We are currently seeking a Research Fellow in Eco-epidemiology/Human Ecology to join our team at Flinders University.
The successful candidate will develop spatial eco-epidemiological models for the populations of Indigenous Australians exposed to novel diseases upon contact with the first European settlers in the 18th Century. The candidate will focus on:
developing code to model how various diseases spread through and modified the demography of the Indigenous population after first contact with Europeans;
contributing to the research project by working collaboratively with the research team to deliver key project milestones;
independently contributing to ethical, high-quality, and innovative research and evaluation through activities such as scholarship, publishing in recognised, high-quality journals and assisting the preparation and submission of bids for external research funding; and
supervising of Honours and postgraduate research projects.
The ideal candidate will have advanced capacity to develop eco-epidemiological models that expand on the extensive human demographic models already developed under the auspices of the Australian Research Council Centre of Excellence for Australian Biodiversity and Heritage, of which Flinders is the Modelling Node. To be successful in this role, the candidate will demonstrate experience in coding advanced spatial models including demography, epidemiology, and ecology. The successful candidate will also demonstrate:
Wildfires transform forests into mosaics of vegetation. What, where, and which plants thrive depends on when and how severely a fire affects different areas of a forest. Such heterogeneity in the landscape is essential for animal species that benefit from fire like woodpeckers.
The black-backed woodpecker (Picoides arcticus) lives in the coniferous forests of North America’s boreal-Mediterranean region. Thanks to a powerful and sharp bill, this bird can excavate nests inside the trunks of (mainly dead) trees, and those cavities will be re-used later by many species of birds, mammals, and invertebrates in fire-prone landscapes (22). The images show a male with the characteristic black plumage of his back that serves as camouflage against the dark bark of a dead tree three years after a wildfire in Montana (USA). Being omnivores, the diet of this bird largely relies on the larvae of woodboring coleoptera like jewell and longhorn beetles. These insects are abundant post-fire, the champion being the fire beetle (Melanophila spp.). The thorax of fire beetles is equipped with infrared-light receptors that can detect a wildfire from tens of kilometres away (23). These fascinating little beasts are the first to arrive at a burned forest and, of course, woodpeckers follow soon after. The preference of the blackbacked woodpecker for burned forests and their cryptic feathers and pyrophilic diet reflect a long evolutionary history in response to fires. Courtesy of Richard Hutto.
Anyone raised in rural areas will have vivid recollections of wildfires: the thick, ashy smell, the overcast sky on a sunny day, and the purring of aerial firefighters dropping water from their hanging tanks. The reality is that wildfires are natural events that shape biodiversity and ecosystem function (1) — to the extent that fire is intimately linked to the appearance and evolution of terrestrial plants (2). Since the Palaeolithic, our own species has used fire at will, to cook, hunt, melt metals, open cropland or paths, or tell stories in front of a hearth (3).
Where there are regular wildfires (fire-prone ecosystems), different areas of the landscape burn in different seasons and years under different weather patterns. Therefore, each region has a unique fire biography in terms of how frequently, how much, and how long ago wildfires occurred. All those factors interact will one another and with topography.
Thanks to the collaborative and evidence-driven foresight of my colleagues at PIRSA Biosecurity and Landscape Boards, I was recently involved in more research examining the most efficient, cost-effective, and humane ways to cull feral dear in South Australia. The resulting paper is now in review in NeoBiota, but we have also posted a pre-print of the article.
Feral deer are a real problem in Australia, and South Australia is no exception. With six species of feral deer in the country already (fallowDama dama, redCervus elaphus, hogAxis porcinus, chitalA. axis), rusaC. timorensis, and sambarRusa unicolor deer), fallow deer are the most abundant and widespread. These species are responsible for severe damage to native plants, competition with native animals, economic losses to primary industries (crops, pastures, horticulture, plantations), and human safety risks from vehicle collisions. Feral deer are also reservoirs and vectors of endemic animal diseases and have the potential to transmit exotic animal diseases such as foot-and-mouth. If left uncontrolled, within 30 years the economic impacts of feral deer could reach billions of dollars annually.
We’ve just published a paper in PLOS ONE showing high infant mortality rates are contributing to an incessant rise of the global human population, which supports arguments for greater access to contraception and family planning in low- and middle-income nations.
In the first study of its kind, we provide a compelling argument that the United Nations’ Sustainable Development Goals for reducing infant mortality can be accelerated by increasing access to family planning.
Although it sounds counterintuitive, higher baby death rates are linked to higher population growth because the more babies a women loses, the more children she is likely to have. Family planning, including access to quality contraception, enables women to plan pregnancies better and therefore reduce infant mortality to curb the so-called ‘replacement’, or ‘insurance’ effect.
We evaluated six conditions thought to influence a woman’s fertility — availability of family planning, quality of family planning, education, religion, mortality, and socio-economic conditions, across 64 low- to middle-income countries.
Not exactly a conservation topic, I know, but it does provide insights into how the ancestors of Indigenous Australians adapted to and thrived in a new and sometimes harsh landscape. The more I study elements of human ecology in deep time, the more awed I become at the frankly amazing capacity of First Peoples.
We combined new models of demography and wayfinding based on geographic inference to show the scale of the challenges faced by the ancestors of Indigenous people making their mass migration across the supercontinent more than 60,000 years ago.
The ancestors of Aboriginal people likely first entered the continent 75,000–50,000 years ago from what is today the island of Timor, followed by later migrations through the western regions of New Guinea.
This pattern led to a rapid expansion both southward toward the Great Australian Bight, and northward from the Kimberley region to settle all parts of New Guinea and, later, the southwest and southeast of Australia.
We did this research under the auspices of the ARCCentre of Excellence for Australian Biodiversity and Heritage (CABAH) and including international experts in Australia and the United States to investigate the most likely pathways and the timeframe needed to reach population sizes able to withstand the rigours of their new environment.
By combining two existing models predicting the routes these First Peoples took – ‘superhighways’ – and the demographic structure of these first populations, we were able to estimate the time for continental saturation more precisely. The new research has just been published in the journal Quaternary Science Reviews.
Based on detailed reconstructions of the topography of the ancient continent and models of past climate, we developed a virtual continent and programmed populations to survive in and move successfully through their new territory.
Navigating by following landscape features like mountains and hills and knowing where to find water led to successful navigation strategies. The First Peoples of Australia soon passed along cultural knowledge to subsequent generations facilitating the peopling of the whole continent.
Each organism has a limit of tolerance to cold and hot temperatures. So, the closer it lives to those limits, the higher the chances of experiencing thermal stress and potentially dying. In our recent paper, we revise gaps in the knowledge of tolerance to high temperatures in cold-blooded animals (ectotherms), a diverse group mostly including amphibians and reptiles (> 16,000 species), fish (> 34,000 species), and invertebrates (> 1,200,000 species).
As a scientist, little is more self-realising than to write and publish a conceptual paper that frames the findings of your own previous applied-research papers. This is the case with an opinion piece we have just published in Basic and Applied Ecology1 — 10 years, 4 research papers2-5 [see related blog posts here, here, here and here], and 1 popular-science article6 after I joined the Department of Biogeography and Global Change (Spanish National Research Council) to study the thermal physiology of Iberian lizards under the supervision of Miguel Araújo and David Vieites.
Iberian lizards for which heat tolerance is known (varying from 40 to 45 °C)
[left, top to bottom] Iberian emerald lizard (Lacerta schreiberi, from Alameda del Valle/Madrid) and Geniez’s wall lizard (Podarcis virescens, Fuertescusa/Cuenca), and [right, top to bottom] Algerian sand racer (Psammodromus algirus, Navacerrada/Madrid), Andalusian wall lizard (Podarcis vaucheri, La Barrosa/Cádiz), Valverde’s lizard (Algyroides marchi, Riópar/Albacete), and Cyren’s rock lizard (Iberolacerta cyreni, Valdesquí/Madrid). Heat-tolerance data deposited here and used to evaluate instraspecific variation of heat tolerance3,4. Photos: Salvador Herrando-Pérez.
In our new paper, we examine how much we know and what areas of research require further development to advance our understanding of how and why the tolerance of ectotherm fauna to high environmental temperature (‘heat tolerance’ hereafter) varies within and across the Earth’s biomes. We focus on data gaps using the global database GlobTherm as a reference template (see Box 1 below).
Our three main tenets
1. Population versus species data: Most large-scale ecophysiological research is based on modelling one measurement of heat tolerance per species (typically representing one population and/or physiological assay) over hundreds to thousands of species covering broad geographical, phylogenetic, and climatic gradients.
But there is ample evidence that heat tolerance changes a lot among populations occupying different areas of the distribution of a species, and such variation must be taken into account to improve our predictions of how species might respond to environmental change and face extinction.
The Faculty for Mathematics and Natural Sciences of Humboldt-Universität zu Berlin (HU Berlin), Geography Department, has an open position for a tenure-track professorship in Conservation and Development.
Starting as soon as possible. This is a Junior Professorship (W1 level, 100%) with a tenure track to a permanent professorship (W2 level, 100%). To verify whether the individual performance meets the requirements for permanent employment, an evaluation process will be opened not later than four years of the Junior Professorship. Tenure track professors at the HU Berlin are expected to do research and teaching, as well as to be active in university administration, in the promotion of young scientists, and in acquiring leadership and management skills. The concrete requirements out of the framework catalogue will be specified in the course of the appointment process.
We seek candidates with an outstanding research record in biodiversity conservation and sustainable development, with experience in working in the Global South. Successful candidates are rooted in conservation science and must have a doctoral degree in conservation science, development geography, environmental science, political ecology or related fields. We expect a demonstrated ability to work interdisciplinary, across the social and natural sciences to understand conservation challenges and and develop solutions.
We seek individuals with the vision, leadership and enthusiasm to build an internationally recognised research program. We expect collaboration with other research groups at the department, at HU Berlin and beyond, and a commitment to promoting a positive, diverse, and inclusive institutional culture. Experience in translating conservation science into action and/or work at the science/policy interface are beneficial.
We offer a tenure-track position in an international, young and vibrant department with an excellent scientific and education track record. The successful candidate will join an interdisciplinary group of faculty focused on human-environment relations, global change, and sustainability.
The salary will be according to W1 level, and after successful tenure evaluation W2 level. Employment at HU Berlin offers all benefits of the German public service system, including health insurance, an attractive pension plan, and social benefits.
Flooding in the Murray-Darling Basin is creating ideal breeding conditions for many native species that have evolved to take advantage of temporary flood conditions. Led by PhD candidate Rupert Mathwin, our team developed virtual models of the Murray River to reveal a crucial link between natural flooding and the extinction risk of endangered southern bell frogs (Litoria raniformis; also known as growling grass frogs).
Southern bell frogs are one of Australia’s 100 Priority Threatened Species. This endangered frog breeds during spring and summer when water levels increase in their wetlands. However, the natural flooding patterns in Australia’s largest river system have been negatively impacted by expansive river regulation that some years, sees up to 60% of river water extracted for human use.
Our latest paper describes how we built computer simulations of Murray-Darling Basin wetlands filled with simulated southern bell frogs. By changing the simulation from natural to regulated conditions, we showed that modern conditions dramatically increase the extinction risk of these beloved frogs.
The data clearly indicate that successive dry years raise the probability of local extinction, and these effects are strongest in smaller wetlands. Larger wetlands and those with more frequent inundation are less prone to these effects, although they are not immune to them entirely. The models present a warning — we have greatly modified the way the river behaves, and the modern river cannot support the long-term survival of southern bell frogs.’
Following my annual tradition, I present the retrospective list of the ‘top’ 20 influential papers of 2022 as assessed by experts in Faculty Opinions(formerly known as F1000). These are in no particular order. See previous years’ lists here: 2021, 2020, 2019, 2018, 2017, 2016, 2015, 2014, and 2013.
Climate change is one of the main drivers of species loss globally. We know more plants and animals will die as heatwaves, bushfires, droughts and other natural disasters worsen.
But to date, science has vastly underestimated the true toll climate change and habitat destruction will have on biodiversity. That’s because it has largely neglected to consider the extent of “co-extinctions”: when species go extinct because other species on which they depend die out.
Our new research shows 10% of land animals could disappear from particular geographic areas by 2050, and almost 30% by 2100. This is more than double previous predictions. It means children born today who live to their 70s will witness literally thousands of animals disappear in their lifetime, from lizards and frogs to iconic mammals such as elephants and koalas.
But if we manage to dramatically reduce carbon emissions globally, we could save thousands of species from local extinction this century alone.
Ravages of drought will only worsen in coming decades. CJA Bradshaw
An extinction crisis unfolding
Every species depends on others in some way. So when a species dies out, the repercussions can ripple through an ecosystem.
For example, consider what happens when a species goes extinct due to a disturbance such as habitat loss. This is known as a “primary” extinction. It can then mean a predator loses its prey, a parasite loses its host or a flowering plant loses its pollinators.
A real-life example of a co-extinction that could occur soon is the potential loss of the critically endangered mountain pygmy possum (Burramys parvus) in Australia. Drought, habitat loss, and other pressures have caused the rapid decline of its primary prey, the bogong moth (Agrotis infusa).
My definition of a ‘lab’ is simply a group of people who do the science in question — and people are a varied bunch, indeed. But I wager that most scientists would not necessarily give much dedicated thought to the diversity of the people in their lab, and instead probably focus more on obtaining the most qualified and cleverest people for the jobs that need doing.
For example, I have yet to meet an overtly racist, sexist, or homophobic scientist involved actively in research today (although unfortunately, I am sure some do still exist), so I doubt that lab heads consciously avoid certain types of people when hiring or taking on new students as they once did. The problem here is not that scientists tend to exclude certain types of people deliberately based on negative stereotypes; rather, it concerns more the subconscious biases that might lurk within, and about which unfortunately most of us are blissfully unaware. But all scientists must be aware of, and seek to address, their hidden biases.
It is time to place my cards on the table: I am a middle-aged, Caucasian, male scientist who has lived in socially inclusive and economically fortunate countries his entire life. As such, I am the quintessential golden child of scientific opportunity, and I am therefore also one of the biggest impediments to human diversity in science. I am not able to change my status per se; however, I can change how I perceive, acknowledge, and act to address my biases.
The earlier scientists recognise these challenges in their career, the more effective they will be.
I acknowledge that as a man, I am already on thin ice discussing gender inequality in science today, for it is a massive topic that many, far more qualified people are tackling. But being of the male flavour means that I have to, like an alcoholic, admit that I have a problem, and then take steps to resolve that problem. After all, privilege is generally invisible to those who have it. If you are a male scientist reading this now, then my discussion is most pertinent to you. If you are female, then perhaps you can use some of these pointers to educate your male colleagues and students.
There is now ample evidence that science as a discipline is just as biased against women as most other sectors of professional employment, even though things have improved since the bad old days of scientific old-boys’ clubs. Journals tend to appoint more men than women on their editorial boards, and that editors display what is known as homophily when selecting reviewers for manuscripts: the tendency to select reviewers of the same gender as themselves.
Likewise, experimental evidence demonstrates that scientists in general rate male-authored science writing higher than female-authored works, and that academic scientists tend to favour male applicants over females for student positions. In the United Kingdom, as I suspect is more or less the case almost everywhere else, female academics in science, engineering, and mathematics also tend to have more administrative duties, and hence, less time to do research; they also have fewer opportunities for career development and training, as well as earning a lower salary, holding fewer senior roles, and being less likely to be granted permanent positions.
Those of us living with cats share our homes with an ancestral predator, one adapted for hunting and the frequent, exclusive consumption of meat. These instincts become fully activated outside the domestic environment, where cats pose a global threat to wildlife. Pets are family. We celebrate their arrival with the same joy as a grand…
Many animals avoid contact with people. In protected areas of the African savanna, mammals flee more intensely upon hearing human conversations than when they hear lions or sounds associated with hunting. This fear of humans affects how species use and move in their habitat. Throughout our lives, we interact with hundreds of wildlife species without…
Deep-sea sharks include some of the longest-lived vertebrates known. The record holder is the Greenland shark, with a recently estimated maximum age of nearly 400 years. Their slow life cycle makes them vulnerable to fisheries. Humans rarely live longer than 100 years. But many other animals and plants can live for several centuries or even millennia, particularly…