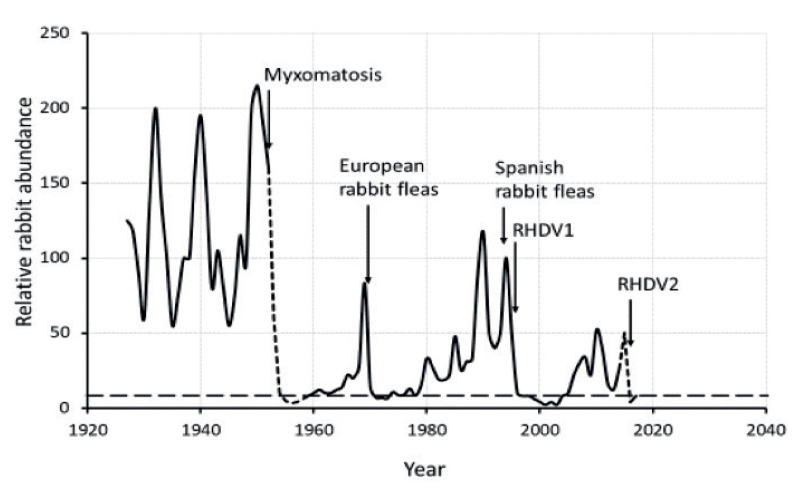
(originally published on the GE.blog)
(originally published on the GE.blog)
Scientists appear to have mixed feelings when it comes to interdisciplinarity in science — the reaction spans from genuine enthusiasm right through to pure disdain.
I myself have crossed many research fields since my Masters project, but despite the support of my supervisors, I have already had to face some tough gatekeeping from science specialists in conferences and in front of other panels. Several times I was taken aback by some reactions, so I have started to become interested in the topic from a more analytical perspective. How are these fields’ boundaries defined in science?
Although each field’s specific methodology, jargon, and tendency to interpret results could represent communication barriers among them, this can be easily overcome by spending time learning the language of other groups, in the company of specialist collaborators, or by attending workshops.
But what about ideology — a philosophy of science inherent to a specific group of individuals? This is one of the things making us human. It definitely affects our society, and even if it is never assumed, it also affects the generation of scientific knowledge from its production to its transmission. Scientists have that connection to their field, its history, its identity, and its compromises.
For example, historians or philosophers use different ways of thinking than do physicists or biologists. The first group aims to clarify and analyse the reconstruction of past events, while the second group strives for conceptual understanding. While useful withina field, these specific ways of seeing science can generate roadblocks when two fields need to start a conversation.
I will tell you a story based on my own experience. Read the rest of this entry »
 We are most fortunate that Dr
We are most fortunate that Dr 


 So, we go through the motions; we
So, we go through the motions; we  Just under two weeks ago,
Just under two weeks ago, 



 This might seem a little left-of-centre for
This might seem a little left-of-centre for  So, a few of us have just submitted a letter contesting the Western Australia Government’s recent decision to delist dingoes as ‘fauna’ (I know — what the hell else could they be?). The letter was organised brilliantly by Dr
So, a few of us have just submitted a letter contesting the Western Australia Government’s recent decision to delist dingoes as ‘fauna’ (I know — what the hell else could they be?). The letter was organised brilliantly by Dr  Last week we had the pleasure of welcoming the eminent sustainability scientist, Professor
Last week we had the pleasure of welcoming the eminent sustainability scientist, Professor